농업직, 농촌지도사, 농업연구사, 방통대 시험대비 꼭 알아야 할 작물생리학 핵심 요약정리! 5장 호흡
5장 호흡
<호흡작용-respiration>
: 식물은 체외에서 산소를 흡수하여 체내의 유기물질인 탄수화물, 단백질 또는 지방을 산화하여 생육에 필요한 에너지를 유리시키고 체외로 탄산가스와 물을 배출
* 호흡 과정 : C6H12O6 + 6O2 + 6H2O → 6CO2 + 12H2O + 36ATP
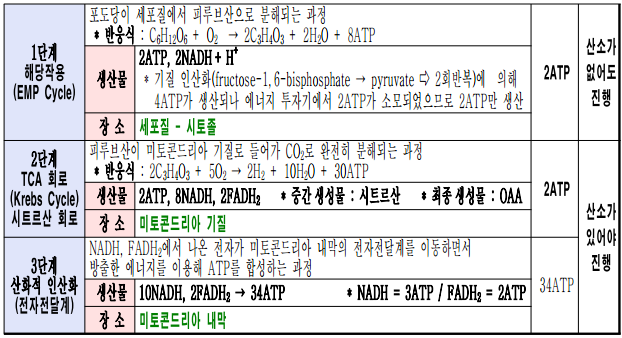
* 호흡작용의 과정(4단계) : 해당작용 - acetyl-CoA 형성 - TCA회로 - 전자전달계
☞ 포도당 1분자가 세포호흡에 의하여 CO2 + H2O 물로 완전히 분해될 때 총 38ATP가 생산되나 해당과정에서 생산된 2NADH가 미토콘드리아로 이동을 하면서 2ATP를 소모하게 되어 최종적으로는 36ATP가 생성된다.
☞ 호흡작용의 4단계 : 해당작용(EMP) → Acetyl CoA 형성 → TCA회로 → 전자전달경로
☞ Acetyl CoA 형성에 필요한 5종의 보조인자 : thiamine pyrophosphate(TPP), Mg이온, NAD+, coenzyme A, 리포산
* 5탄당 인산회로(6탄당인산분지회로) : glucose - 6 – phosphate로부터 시작(탈수소반응)하여 포도당을 산화하는 대사회로
- 해당과정과 유사하여 공통의 반응물을 가지며, 주로 세포질에서 일어난다.
※ 전자수용체 : 5탄당 인산회로 – NADP+/ 해당과정 – NAD+
- glucose - 6– phosphate 1분자가 5탄당인산회로를 통하여 산화될 때에는 두 반응에서 2분자의 NADPH+H +가 생성되며 CO2와 물로 완전히 산화된다면(6회의 회로) 12 NADPH+H+가 생성된다.
- 세포내에서 5탄당인산회로를 통하여 NADPH+H+는 일반적으로 전자전달계에 의하여 ATP로 산화되지 않고 NADPH+H+를 전자공여체로서 특별히 요구하는 지방산과 isoprenoid의 합성과 같은 합성반응에 더 많이 이용
- 생장 중인 어린 조직에서는 포도당 산화의 주된 회로로서 크렙스회로를 이용하지만, 식물의 지상부와 늙은 조직에서는 크렙스회로뿐만 아니라 5탄당인산회로도 이용, 체내에서 포도당대사의 약 10%는 이 5탄당인산회로를 통하여 이루어진다
<에너지 운용>

* 포도당 1분자의 에너지 효율(1ATP = 7.3Kcal) : 1C6H12O6
= 686Kcal ⇨ 산소 호흡시 발생 36ATP(262.8Kcal)
⇨ 에너지 효율 40% / 발열-호흡열 60%
* TCA(크렙스) 회로의 최종산물 : 옥살초산 ☞ 시트르산 (구연산 ) → α -케토글루타르산 → 숙신산 → 푸마르산 → 말산 → 옥 살 아 세트산
* 광합성율은 온도가 상승함에 따라 호흡률보다 서서히 증가
* 광호흡(Photorespiration)현상 - peroxisome이라는 특정 기관에서 일어난다.
① 탄소 고정량의 감소 유발 ② 광합성 효율 저하 ③ RuBP - Oxygenase가 관여
④ 산소에 의한 광합성 저해 - Warburg Effect ⑤ 산화적 광파괴로부터 보호
↳ * 광 산화는 특히 산 소 가 있는 상 태 에서 더 욱 심하 게 엽 록 소 파 괴
* 광호흡 에 대한 조건 (C3 plant)
① RuBP가 CO2결합 대신 O2와 결합 ② O2의 농도가 높고 CO2의 농도가 낮은 조건 ③ 광도하에 반응, 고온에서 촉진
* 온도에 따른 광합성 능력비교
일반적으로
- 고온조건에서는 C4작물이 C3작물보다
- 저온조건에서는 C3작물이 C4작물보다 높다
* 지방의 호흡계수 : 지방이 당으로 변화할 때 산소를 흡수하나 이산화탄소를 방출하지 않아서 호흡계수는 0이다.
* 6탄당이 호흡작용에 의하여 산화되는 경우 실제로 호흡원으로서 이용되는 것은 당류 자체가 아니고 인산화된 당류인 fructose - 1, 6 - bisphosphate이다.
* 세포막에 존재하고 있는 양이온펌프를 작동하여 H+를 세포질 외부로 내보내서 세포질을 전기적으로 음성으로 만들기 위해서는 ATPase에 의하여 ATP가 분해 되어야한다.
* 산소가 충분히 존재할 때 해당과정에 의하여 생성된 피루브산은 산화적으로 탈탄산화되어 acetyl CoA가 된다.
→ acetyl CoA는 해당과정와 크렙스회로를 연결시키는 역할을 한다.
* 알코올 발효는 해당과정에서 생긴 피루브산이 탈탄산작용에 의하여 아세트알데하이드가 되고 알코올탈수소효소에 의하여 알코올로 환원된다.
* 호흡작용의 최저온도에서 최적온도에 이를 때까지의 사이에는 온도가 10℃ 상승할 때마다 호흡률은 약 2배가 된다.
* 어떤 온도에 있어서의 반응속도가 이보다 10℃높은 온도에서 몇배로 되는가를 나타내는 값을 Q10 즉 온도계수라고 한다.
- 논벼에서 호흡작용의 Q10은 일반적으로 1.6~2.0이다.
* 호흡작용 : 대기중의 CO2 농도가 높아지면 일반적으로 호흡속도는 감소하나 예외도 있다.
→ 온도가 낮고, O2가 부족할 때, 고농도 CO2에서 호흡저하는 더욱 현저하다. CO2에 의한 호흡억제는 과실이나 채소의 저장에 이용된다. 예) CA저장

* 질소반응 부정적 작물(질소에 대해 부(負) 의 방향으로 개화 반응 하는 작물)
- 벼, 밀, 귀리, 화본과목초, 알팔파, 붉은토끼풀 - 소식하면 출수개화 늦어지므로 밀식해야한다
※ 다량의 질소가 탄질비 저하시켜 질소반응이 부의 작물에 개화를 지연시킨다.
※ 소식을 하게 되면 1개체당 질소량이 많아지기 때문이다.
☆ 질소가 작물의 성숙과 수량에 비치는 영향
- 보리, 밀, 귀리, 호밀 등 온대성 C3 화곡류는 질소함량이 높으면 경엽의 생장이 지나쳐서 성숙이 지연되고 조고비율 (종실의 짚에 대한 비율)이 낮아지며, 도복하기 쉽다.
- 한편 옥수수, 수수 등 열대성 C4 화곡류는 질소함량이 높으면 개화 및 성숙이 빨라지고 질소과잉의 해는 적다.
벼의 성숙과 조고비율에 대한 질소의 영향은 온대성 화곡류형과 열대성 화곡류형의 중간이다.
- 벼를 무질소로 재배하면 기본영양생장이 억제되어 출수가 지연되지만, 질소를 적량보다 많이 시용하면 영양생장이 과도하게 촉진되어 간장이 길어지고, 특히 절간신장기에는 하위절간이 신장되어 도복하기 쉬우며, 출수가 다소 지연되고, 도열병에 쉽게 걸린다.

* celluose의 분해
* 단당류의 역할
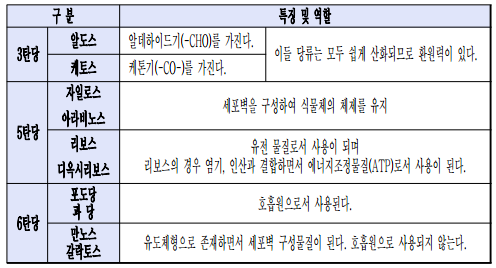
* 스타키오스 : 일부 식물에서 자당 대신에 체내를 전류하는 중요한 탄수화물이 된다.
* 라피노스 : 여러 식물의 잎에 소량 함유되어 있지만 종자와 같은 저장기관에 다량 함유되어 있으며 발아할 때 소모된다.
- 종자형성의 경우와 같이 식물조직에 의한 수분의 상실은 라피노스의 합성을 촉진한다.
* 셀룰로스 : 식물의 1차 세포벽에는 20%, 목재섬유의 2차 세포벽에는 43%, 목화는 90%가 함유되어 있다.
* 탄수화물의 합성과 분해
- 탄수화물의 변형에 있어서 가장 보편적이고 필수적인 특징은 인산화
- 고등식물에 있어서 자당의 생합성은 uridine diphosphate glucose(UDPG)가 관여
* 탄수화물대사
- 엽록체에서 시트르산회로의 생성물로부터 전분이 생성되려면 자당형성에서 요구되는 바와 같이 먼저
glucose-1-phosphate가 형성되어야 한다.
* 전분 합성 경로(세포질에서 합성)
triose phosphate → fructose-1, 6-bisphosphate → glucose-6-phosphate
→ glucose-1-phosphate + ATP → ADP-glucose → 전분
* 환원-비환원당
- 포도당, 과당 등의 단당류는 환원당이지만 설탕(서당, 자당)은 비환원당이다. 비환원당은 환원당에 비해 다른 물질과 화학반응을 일으킬 가능성이 작고 안정적이다. 따라서 체관을 따라 이동할 때에는 반응성이 작고 안정적형태인 설탕의 형태로 이동한다.
* 당류의 전류
- 단당류는 환원력이 있으며 전류가 되지 않음. 세포구성, 호흡원으로 사용됨.
- 다당류(2, 3, 4당류)는 환원력이 없으며 전류가 됨. 저장물질이 됨.
* 대사물질의 체관부 전류 속도 : sucrose(107cm/h) < H 2O, P(87cm/h)
* 전류물질의 운반과 저장
- 식물 호르몬은 수용부 세포의 활성에 영향을 미쳐 동화물질의 분배에 큰 효과를 나타낸다.
- 옥신, 시토키닌, 에틸렌, 지베렐린은 줄기 절단면에 처리하면 처리한 부위에 동화물질의 축적이 유발
* 체관요소는 하나 이상의 반세포와 짝을 이루어 형성되며, 원형질 연락사가 체관요소와 반세포를 관통하여 용질을 쉽게 이동시킨다.
* 유조직세포는 대사적인 펌프로서 작용하여 양분을 공급부위에 있는 체관부에서 수용부위에 있는 체관부로 보낼 때 필요한 에너지를 공급한다.
* 수용부위 말단의 하적은 팽압을 낮은 상태로 유지하도록 하고 당의 농도를 낮게 하며 물이 빠져 나오게 하고 공급원으로 부터 압력을 전달받을 수 있게 한다.
* 생식기관 수용부위(발육중인 종자)에서는 배조직과 모계조직 간에 심플라스트로 연결되지 않기 때문에
아포플라스트 단계가 필요하다.
* 동화물질의 분배
- 수용부위의 동화물질의 수용능력인 수용부 강도는 수용부의 크기와 활성 두 요인에 의하여 좌우된다.
수용부 강도 = 수용부 크기 × 수용부 활성
- 수용부 크기는 수용부 조직의 전체 무게이고 수용부 활성은 수용부 단위중량당 동화물질의 흡수속도이다.
- 식물호르몬은 수용부 세포의 활성에 영향을 미쳐 동화물질의 분배에 큰 효과를 나타낸다.
인돌초산, 시토키닌, 에틸렌 및 지베렐린을 줄기 절단면에 처리하면 부위에 동화물질의 축적이 유발된다.
* 양분저장과 환경조건
- 동화물질의 저장에는 여러 환경조건이 영향을 미친다. 작물의 생장상태는 앞에서 설명한 바와 같이 양분의 저장에 영향을 주며 생장이 왕성해지면 동화물질은 새로운 조직의 형성이나 호흡작용에 소비되는 일이 많고 저장양분의 축적이 적어지거나 또는 저장되어 있는 양분의 소모를 가져오는데 생장이 감퇴되면 이러한 소모가 적고 저장이 많아진다.
* 질소화합물 대사
- 세포에 흡수된 질산태 질소는 식물의 질소화합물에 결합되기 전에 에너지를 이용하여 암모늄으로 환원되어야 한다.
- 요소는 식물의 유기화합물 속에 결합되기 이전에 urease의 작용으로 가수분해되어 암모니아와 탄산가스로 전환된다.
* 비공생적 질소고정
- 질소고정세균은 독립생활을 하면서 비공생적 질소고정을 한다.
질소고정세균에는 절대호기성균(Azotobacter), 통성혐기성균(Bacillus), 혐기성균(Clostidium),
광합성세균(Rhodospirillum) 등이 있다.
* 공생적 질소고정
- 기주식물과 세균의 정확한 상호 인지가 필수적이다. 기주식물은 세균의 유전자 발현을 유도하는 플라보노이드 물질을 분비하고 세균은 이에 반응하여 2차 신호로 작용하는 지질올리고당 뿌리혹형성인자를 생성한다.
세균의 침입에는 특이적인 세포 외 다당류가 필요하다.
- 세균에 의해 고정된 질소는 암모늄 형태로 세균상체에서 식물세포의 세포질로 운송되어 글루타민합성효소에 의해 글루타민에 동화된다
* 단백질의 생합성
① 사슬합성의 시작 : 번역개시복합체를 형성 - 메싸이오닌-tRNA + 진핵세포개시인자(eIF) + GTP
↳ 40S : Met-tRNA : 개시인자복합체
복합체가 5’ 말단으로부터 첫 번째 AUG자리를 찾으면 mRNA의 암호와 tRNA의 역암호 사이에
상호작용이 진행된다. 암호와 역암호의 상호작용이 일어나면 대단위체 리보솜이 결합하고
개시인자와 GDP가 유리되어 80S 번역개시복합체를 완성한다.
※ 식물과 다른 진핵세포의 단백질합성과정중 다른점은 식물에는 소단위체 리보솜이 결합기 전에 5’ 말단을
인식하는 두 가지 유형의 개시인자(eIG4F)가 존재한다는 것이다.
② 사슬연장 : 번역복합체가 mRNA를 5’ → 3’ 방향으로 이동하면서 유전암호와 역유전암호 사이의 상호작용을 통하여 첫 번째 아미노산의 메싸이오닌에 다음 아미노산이 연속적으로 첨가되어 폴리펩타이드가 신장되는 과정이다.
③ 사슬종결 : 폴리펩타이드사슬의 신장은 리보솜의 A자리에 3종류의 종결암호(UAA, UAG, UGA) 중의 하나가 위치하면 종결된다. 종결암호에 대응하는 tRNA는 존재하지 않으며 대신 유리인자가 종결암호가 위치한 A자리에 결합하여 종결을 유도한다.
※ 80S 번역개시복합체로 조립된 리보솜에는 아미노아실(A)자리, 펩타이드(P) 및 출구(E)자리가 있다.
* 피리미딘 뉴클레오티드 생합성장소 : 식물에서는 피리미딘이 주로 색소체에서 합성되며, 생합성에 관여하는 효소가 모두 색소체에 존재한다.
* 뉴클레오티드의 대사
- 식물체 또는 세포는 대상과정의 요구도에 따라 뉴클레오티드의 총량을
① 새로운 합성, ② 분해, ③ 회수(재활용), ④ 인산전이반응 등의 경로를 통하여 조절한다.
* 뉴클레오티드의 세포내 분포
- 식물세포에는 아데닌과 유리딘 뉴클레오티드가 구아닌과 시토신 뉴클레오티드보다 훨씬 많이 존재하는데
이는 아데닌과 유리딘 뉴클레오티드가 핵산의 합성과 대사의 보조인자로서 더 광범위하게 이용되기 때문이다.
☞ 식물세포 내 분포 : A, U > G, C
- Adenine nucleotide 일반적 존재 : 세포질 46%, 색소체 45%, 미토콘드리아 9%
- ATP / ADP 비율 : 세포질 > 빛 엽록체 > 암 염록체
* 단백질의 분해
- 세포 내 단백질의 약 절반은 매 4~7일마다 교환된다. 종자가 발아할 때에는 단백질 분해효소가 현저하게 증가하고, 이들 효소가 저장단백질을 가수분해하여 생긴 아미노산을 배에 공급하는 역할을 한다.
* 단백질의 구조
- 단백질의 전체적인 모양은 기본적으로 아미노산 잔기에 결합된 원자나 원자단의 입체적 배열, 펩타이드 결합의 이중결합적 성질, 그리고 아미노산 잔기 간의 상호작용에 관여하는 비공유결합적 인력과 이황화결합 등에 의해 결정된다.
* 광합성과 세포 호흡의 전과정 비교
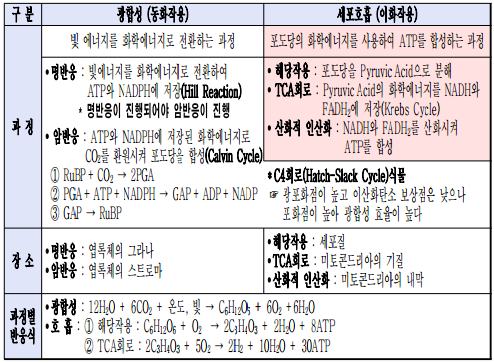
* 광합성과 세포 호흡의 전과정 비교
* 호흡기질 : 세포호흡을 통해 분해되어 에너지를 방출하는 유기물(탄수화물, 지방, 단백질)
* 호흡원의 양 : 당류, 전분 등의 탄수화물이 체내에 많아지면 호흡이 증가하고 이들이 줄어들면 호흡이 저하되고 생육중인 작물체의 체내 당류가 감소하면 호흡작용이 약해진다

* 호흡 기질이 세포 호흡에 이용되는 경우
* 호흡률(CO 2 / O2) : 호흡 기질이 세포 호흡에 의해 분해될 때, 소비된 O2의 부피에 대해 발생한 CO2의 부피의 비
|
∙ 호흡률(RQ) = 발생한 CO2의 부피(CO2 방출량) / 소비된 O2의 부피(O2 흡수량)
∙호흡률은 호흡 기질의 탄소, 수소, 산소의 함량비에 따라 다르게 나타난다.
* 탄수화물의 호흡률 : 1.0, 단백질의 호흡률 : 0.8, 지방의 호흡률 : 약 0.7
⇨ 지방은 당과 비교하여 산소가 적고 수소가 많기 때문에 산화되어 CO2와 H2O가 되기 위해서는 더 많은 산소가
필요하게 되어 호흡계수는 1보다 작다.
|
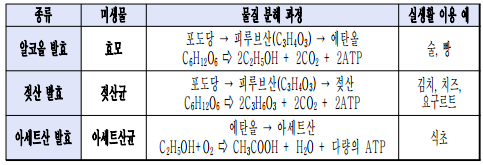
* 발 효
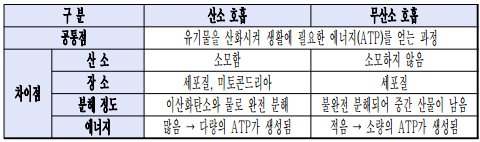
- 산소 호흡과 무산소 호흡의 비교
- 발효의 종류 : 중간 산물의 종류에 따라 알코올, 젖산, 아세트산 발효
* 호흡작용에 미치는 영향
O2의 농도 : 공기 20% 이하면 호흡작용 저하, 5% 이하면 유기호흡 현저히 감소, 발아하는 종자는 산소농도가 낮을 때 유기호흡보다 무기호흡을 더 많이 하나 산소농도가 10% 이상이면 유기호흡을 한다.
온 도 : 식물의 호흡작용은 효소에 의하여 일어나기 때문에 온도의 영향을 받는다.
CO2 농도 : 보통의 CO2농도보다 더 높은 농도에서 호흡작용이 상당히 감소, 기공이 닫혀 산소량 감소하기 때문이다.
수 분 : 호흡율이 매우 낮은 성숙한 건조종자나 포자에 있어서 수분은 호흡의 제한요인이 된다.
화학약제 : cyanamide, azide, fluoride등의 약제는 호흡효소의 작용을 억제 / ethylene 등은 호흡을 촉진
※ 부족하지만 글의 내용이 도움이 조금이라도 되셨다면, 단 1초만 부탁드려도 될까요? 로그인이 필요없는 하트♥(공감) 눌러서 블로그 운영에 힘을 부탁드립니다. 그럼 오늘도 행복한 하루 되십시오^^