11. 지질대사
들어가기
⋅우리 몸에서 이용되는 지질들은 식사를 통해 공급 or 체내 합성됨
⋅중성지방 → 에너지의 주요 저장 형태 → 대사를 통해 에너지를 공급
⋅콜레스테롤 → 담즙생성 및 비타민 D 합성에 기여
⋅불포화지방산 → 생체기능 조절하는 물질들을 생성
⋅당지질 & 인지질 → 주요 막 성분을 이룸
1. 지질의 소화·흡수와 이동
1) 지질의 소화
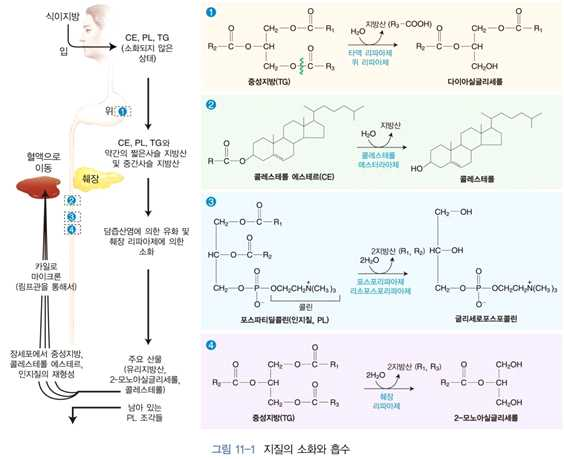
⋅식사를 통해 섭취한 지방의 소화는 위에서부터 시작됨
⋅타액 리파아제 & 위 리파아제 작용
→ 중성지방 10~30%가 위에서 소화됨
→ 위에서 분비되는 리파아제들은 주로 탄소 수 12개 이하의 짧은사슬 또는 중간사슬 지방산을
함유한 중성지방의 소화에 관여함
→ 3번 위치에 있는 지방산을 우선적으로 가수분해함
→ 신생아에서 특히 유지방의 소화 흡수에 중요한 역할을 함
⋅부분적으로 가수분해 된 지방, 중성지방, 콜레스테롤, 인지질 등은 작은 기름방울의 형태로 십
이지장으로 들어오게 됨
⋅담낭에서 분비된 담즙에 있는 담즙산염(bile salt)의 도움으로 유화액 상태를 이룸
→ 유화액 형성은 기름 방울들이 미셀(micelle)의 작은 상태로 안정화될 수 있도록 도와줌
⋅표면적 증가
⋅췌장에서 분비되는 소화효소의 작용을 잘 받을 수 있게 함
⋅ 본격적인 지질의 소화 (췌장 분비 소화효소)
→ 췌장 리파아제, 코리파아제, 콜레스테롤 에스터라아제, 포스포리파아제 A2에 의해 이루어짐
- 췌장 리파아제 : 중성지방
→ 다이아실글리세롤, 모노아실글리세롤 및 유리지방산으로 분해
- 콜레스테롤 에스터라아제: 콜레스테롤 에스테르
→ 유리 콜레스테롤과 지방산으로 가수분해
- 포스포리파아제 A2 : 인지질
→ 리소인지질과 유리지방산으로 분해
2) 지질의 흡수
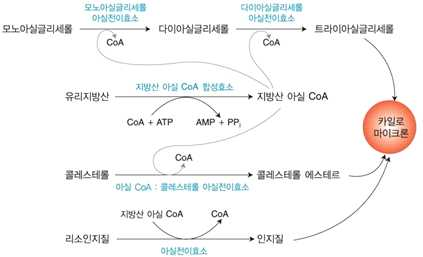
⋅소화된 지방의 흡수 → 주로 소장의 십이지장 말단과 공장부분에서 이루어짐
⋅짧은 사슬 지방산과 중간사슬 지방산은 미셀의 도움 없이도 장 세포에서 흡수될 수 있음
⋅모노아실 글리세롤 → 다이아실글리세롤, 지방산, 콜레스테롤, 리소인지질
→ 미셀 형태로 흡수 → 다시 중성지방, 콜레스테롤에스테르, 인지질로 재합성
→ 트리아실글리세롤 → 카일로마이크론
⋅유리지방산 → 지방산 아실 CoA
⋅콜레스테롤 → 콜레스테롤 에스테르 → 카일로마이크론
⋅리소인지질 → 인지질 → 카일로마이크론
⋅카일로마이크론 림프계를 거쳐 혈액으로 합류
| ※미셀 : 양극성을 가진 지질이 수용액 안에서 소수성인 꼬리는 안쪽을 향하고 친수성인 머리 부분이 수용액 쪽을 향하고 있는 구형의 집합체 |
3) 지단백질의 구조와 종류
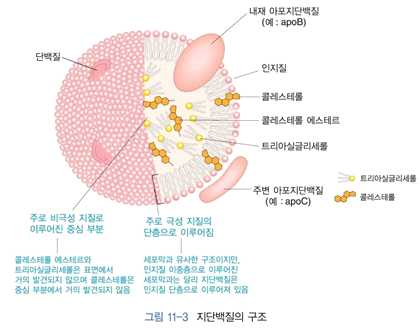
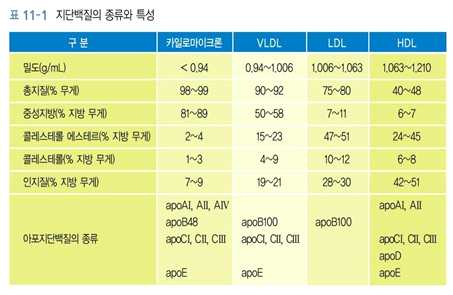
⋅혈액 중에서 소수성인 지질의 주요 이동수단 → 간과 장에서 합성
⋅중심부 : 소수성인 중성지방 + 콜레스테롤 에스테르
⋅표면 : 단층의 인지질 + 에스테르 결합되어 있지 않은 유리 형태의 콜레스테롤 + 아포지단백
질
⋅지단백질의 단백질 부분 → 아포지단백질(apolipoprotein) → 지단백질의 구성, 구조, 대사 및
기능에 중요한 역할을 함
⋅아포지단백질은 수용체의 인식에 기여함
⋅일부 아포지단백질들 → 지질대사에 관여하는 효소들의 활성을 조절함
⋅지단백질은 보통 한 가지 이상의 아포지단백질을 가지고 있음
⋅단백질과 지질의 조성에 따라 지단백질의 밀도가 달라짐
⋅지질의 양 ↑→ 지단백질의 밀도 ↓
⋅카일로마이크론, VLDL이 중성지방이 주요성분
⋅카일로마이크론 → 식사를 통해 섭취한 지방을 장으로부터 조직으로 운반
⋅VLDL → 간에서 합성된 중성지방을 조직으로 운반
4) 지단백질의 대사
(1) 카일로마이크론의 대사
⋅지단백질 중에서 가장 크기가 크고 중성지방이 많음
⋅장에서 만들어지며, 혈중에서는 특히 식사 후에 증가함
⋅보통 식후 30분~3시간 사이 혈중 농도 최고치
⋅지단백질 리파아제(LPL)의 작용을 받아 지방산과 다이아실글리세롤로 분해함
→ 근육이나 지방조직으로 들어감 → 에너지 생성과 중성지방으로 저장
⋅중성지방을 내놓음에 따라 카일로마이크론 크기가 작아짐
⋅중성지방의 양 감소 → 콜레스테롤과 콜레스테롤 에스테르 함량의 비율이 높은 카일로마이크론
잔여물이 됨
⋅간 조직에는 LPL이 없어 카일로마이크론에 있는 중성지방을 제거 못함
⋅카일로마이크론 잔여물에 대한 수용체 존재
→ 카일로마이크론 잔여물을 혈중에서 제거
→ 그 안에 있는 중성지방과 콜레스테롤을 제거하게 됨
(2) VLDL의 대사
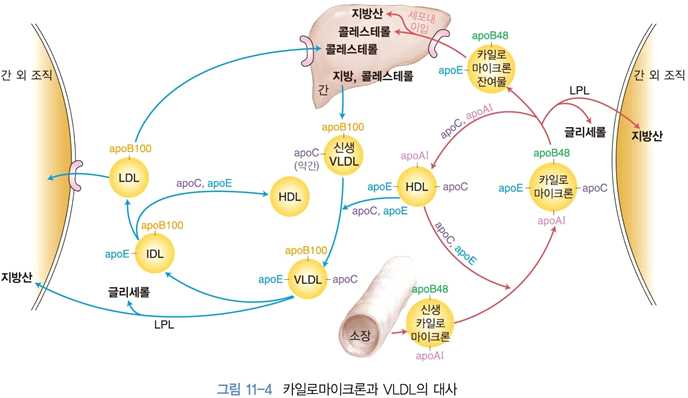
⋅간에서 생성, 체내에서 합성된 중성지방을 조직으로 운반함
⋅VLDL도 LPL의 작용에 의해 내부에 있는 중성지방이 가수분해
→ 근육과 지방조직에 지방산, 다이아실글리세롤을 공급
⋅중성지방 분해 → VLDL 크기 ↓
→ apoC가 떨어져 나감(apoC: LPL 활성에 필요)
⋅중성지방의 양이 줄어든 VLDL의 일부는 간에서 LDL수용체에 의해 제거
(apoE, apoB100이 인식)
⋅일부는 LPL에 의해 중성지방이 더 감소되어 중성지방의 함량은 낮고 콜레스테롤과 콜레스테롤
에스테르가 주요 지질인 LDL로 전환
2. 지방산의 산화
⋅호르몬 민감성 리파아제(HSL) : 지방세포에 저장된 중성지방의 1 또는 3번 탄소에 붙어 있던
지방산이 가수분해 → 유리 지방산의 형태로 분리
⋅에피네프릮, 글루카곤은 HSL을 활성형인 인산화 형태로 전환 → 중성지방의 가수분해 증가 →
혈중 지방산의 농도, 간과 근육조직에서의 지방산 산화 ↑
⋅혈중 인슐린, 포도당 농도 ↑→ 지방산 유리 억제
⋅유리 지방산 : 혈액 내 알부민과 결합 조직으로 이동
⋅지방산 → 세포질에서 지방산 아실 CoA의 형태로 활성화 → 카르니틴에 결합 → 미토콘드리아
기질로 이동
⋅기질내에서 β–산화과정(카르복실기로부터 탄소 두 개 단위로 떨어짐) → 아세틸 CoA를 생성
→ 구연산회로를 통해 산화되면서 에너지를 발생시킴
1) 지방조직으로부터
지방산의 유리 ⋅지방조직에 저장된 중성지방에서 → 지방산이 유리되는 것은 호르몬의 영향을 받음
⋅에피네프린, 글루카곤은 HSL을 활성형인 인산화 형태로 전환 → 지방세포로부터 중성지방의 가
수분해가 증 가하게 함 → 혈중 지방산의 농도, 간과 근육조직에서의 지방산 산화 ↑
⋅혈중 인슐린, 포도당 농도 ↑ → 지방산 유리 억제
⋅유리 지방산은 알부민과 결합되어 혈액 내에서 이동하다가 세포막을 통과하여 세포 내로 들어
오게 됨
2) 지방산의 활성화 및 미토콘드리아 안으로의 이동
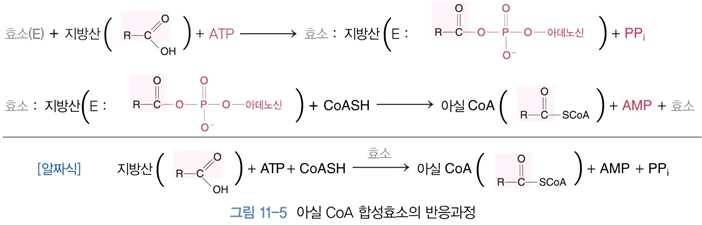
⋅아실 CoA 합성효소는 미토콘드리아의 외막, 소포체, 퍼옥시좀과 결합되어 있으며, 포화지방산
& 불포화지 방산 모두에 작용할 수 있고, 지방산의 사슬 길이에 따라 특이성이 다른 몇 가지
효소가 존재함
⋅세포 내로 들어온 지방산(탄소 10개 이상의 지방산)은 세포질에서 아실 CoA 합성효소에 의해
작용됨
⋅지방산의 카르복실기 + 조효소 A의 황화수소기 사이에 티오에스테르 결합 형성 → 아실 CoA
형태의 활성화된 지방산이 됨
⋅이 반응에서 ATP가 사용되어 AMP와 피로인산으로 전환
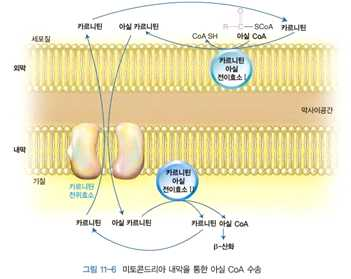
⋅세포질에서 활성화된 지방산 → 미토콘드리아 내막 통과 → β-산화과정에서 의해 대사됨
⋅긴사슬 지방산 아실 CoA는 미토콘드리아 내막을 직접 통과할 수 없음
→ 카르니틴의 도움을 받아 아실카르니틴 형태로 통과
⋅아실 CoA + 카르니틴 아실전이효소 I → 아실 카르니틴으로 전환
⋅탄소 10개 이하의 지방산 → 지방산형태로 진입 → 미토콘드리아 내에서 아실 CoA로 활성화
됨
| ※ 지단백질 리파아제 - 지단백질 안에 있는 중성지방을 지방산과 글리세롤로 분해 ※ 호르몬 민감성 리파아제 -지방세포 안에 있는 중성지방을 지방산과 글리세롤로 분해 -지방세포의 세포질에 있다가 지방방울 표면으로 이동하여 중성지방을 분해 -에피네프린과 글루카곤에 의해 활성 증가 -지방조직에 저장된 중성지방을 분해하여 다른 조직에서 이용될 수 있도록 하는 데 작용함 |
3) 지방산의 산화
(1) 포화지방산의 β-산화
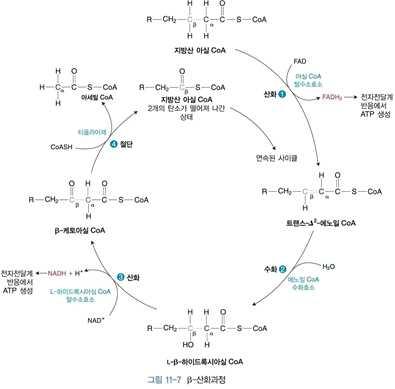
⋅β-산화에서는 4개의 반응을 통해 지방산 아실 CoA의 카르복실기 끝으로부터 탄소 2개가 아세
틸 CoA의 형태로 떨어져 나가고 탄소 2개만큼 짧아진 지방산 아실 CoA는 다시 β-산화를 거
치게 됨
⋅β–산화: (탄소수/2)-1 회
⋅아세틸 CoA : 탄소수/2 개
⋅β–산화 1회 당 : FADH2와 NADH 1개씩 생성함
① 아실 CoA 탈수소효소의 작용에 의한 트랜스 α, β 이중결합의 생성
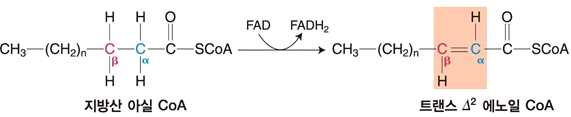
⋅탈수소화 : 트랜스 α, β 이중결합 생성
⋅효소에 결합된 FAD가 FADH2로 환원
⋅생성된 FADH2는 전자전달계에 전달되어 ATP를 생성
② 에노일 CoA 수화효소의 작용에 의한 β-하이드록시아실 CoA 생성
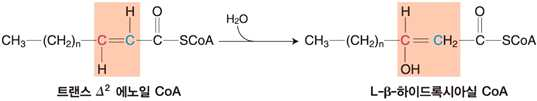
⋅수화 : 이중결합의 β 위치에 -OH 첨가
⋅에노일 CoA 수화효소의 작용에 의해 생성되는 β-하이드록시아실 CoA는 L형
③ L-하이드록시아실 CoA 탈수소효소의 작용에 의한 β-케토아실 CoA의 생성
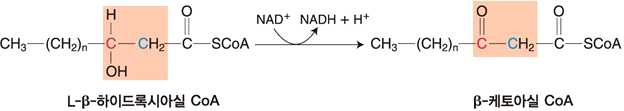
⋅탈수소화 : β 위치에 케토형태 생성
⋅NAD+가 NADH로 환원
⋅생성된 NADH 는 전자전달계에 전달되어 ATP를 생성
④ 티올라아제의 작용에 의한 아세틸 CoA와 탄소 2개가 짧아진 지방산 아실 CoA의 생성

⋅절단 : α, β-탄소 간 결합이 끊어짐
⋅아세틸 CoA가 떨어져 나온 뒤에 CoASH가 티올라아제의 티올기와 치환되면서 지방산 아실
CoA가 형성됨
⋅에노일 CoA 수화효소, 하이드록시아실 CoA 탈수소효소, 티올라아제는 미토콘드리아 내막에 다
중효소복합 체의 형태로 존재함
⋅한 개의 아세틸 CoA는 구연산회로를 통해 1개의 FADH2, 3개의 NADH 및 1개의 GTP를 생성
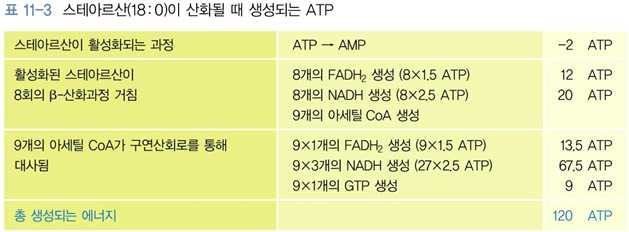
⋅결과적으로 탄소 18개의 탄소를 가진 스테아르산이 산화되면 120 ATP가 생성하게 됨
(2) 불포화지방산의 산화
⋅시스 형태의 이중결합과 이중결합의 위치 때문에 β-산화과정에서 작용하는 에노일 CoA 수화
효소가 작용하지 못함
⋅이성질화를 통해 트랜스 형태로 변환
⋅3번 탄소 시스 형태 → 이성질화 (2번 탄소 트랜스 형태)
⋅4번 탄소 시스 형태 → NADPH 사용 (3번 탄소 트랜스 형태)
(3) 홀수 지방산의 산화
⋅간혹 식물이나 해양 생물에서 생성됨
⋅β-산화되면 마지막에 아세틸 CoA + 프로피오닐 CoA가 생성
⋅탄소수(-3)/2개의 아세틸 CoA와 1개의 프로피오닐 CoA 생성
⋅프로피오닐 CoA → 숙시닐 CoA로 전환되어 구연산회로에서 대사됨
⋅그러나 숙시닐 CoA가 바로 구연산회로에서 대사되는 것이 아님
⋅말산 → 세포질 → 피루브산 → 아세틸 CoA
(4) 퍼옥시좀의 β-산화
⋅탄소 수 22개 이상의 지방산
→ 일차적으로 퍼옥시좀으로 이동 → 매우 긴 사슬 지방산 아실 CoA 합성효소에 의해 활성화된
후 산화
⋅결과적으로 생긴 탄소 수가 적어진 지방산이 미토콘드리아로 이동하여 β-산화가 일어남
⋅퍼옥시좀에 있는 티올라제는 탄소 8개 이하의 아실 CoA에 는 작용핛 수 없음
⋅퍼옥시좀의 산화: FAD가 보조인자로 작용하기는 하나 젂자가 젂자젂달계로 젂달되지 않고 바
로 산소로 젂달되어 과산화수소가 생성 퍼옥시좀 카탈라아제의 작용에 의해 과산화수소는 물과
산소로 전환됨
⋅퍼옥시좀에 있는 티올라제는 탄소 8개 이하의 아실 CoA에는 작용을 하지 못하기 때문에 탄소
수가 작아진 지방산은 미토콘드리아로 이동하여 산화됨
(5) 곁가지 지방산의 α-산화
⋅곁가지가 있는 지방산의 경우 바로 β-산화가 일어나지 못함
⋅퍼옥시좀에서 α-산화와 β-산화를 통해 대사가 이루어짐
(예) 피탄산(phytanic acid) : 클로로필 대사과정에서 생기는 물질
→ 반추동물로부터 얻는 지방과 유제품 등에 존재
⋅피탄산이 산화되면 2-메틸-프로피오닐 CoA, 아세틸 CoA 및 프로피오닐 CoA가 생성됨
4) 지방산 산화의 조절 ⋅혈중 유리지방산의 농도 (HSL에 의해 지방조직에서 유리)에 의해 조절됨
⋅글루카곤 또는 에피네프린 → 지방세포에서 cAMP의 농도 증가 → 단백 질 키나아제 A에 의해
HSL 인산화 → HSL 활성화 → 지방조직으로부터 지방산의 유리 → 혈중 유리지방산 증가 간
과 근육에서 β–산화가 증가
⋅말로닐 CoA (지방산 생합성 과정의 첫 번째 중간대사물)의 농도도 지방산의 산화를 조절함
⋅세포질 내의 말로닐 CoA 농도 증가 → 카르니틴 아실 전이효소 I 작용 억제 → 지방산이 미
토콘드리아 내로 적게 이동 → 결과적으로 지방산의 β–산화 감소
⋅지방산의 합성 & 산화가 동시에 일어나지 않도록 조절함
3. 케톤체의 생성과 이용
⋅케톤체 : 아세토아세트산, β-하이드록시부티르산, 아세톤
⋅간의 미토콘드리아에서 아세틸 CoA로부터 생성됨
⋅혈액 → 말초조직 → 에너지원으로 사용됨
⋅심장과 근육에서 주요 에너지원으로 사용되며, 포도당이 부족할 때는 뇌조직에서도 중요한 에
너지원으로 이용됨
⋅수용성, 혈액 내에서 다른 물질에 결합되지 않은 자유 형태로 이동할 수 있음
1) 케톤체의 생성
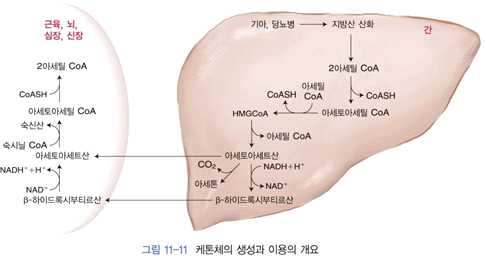
⋅간에서 생성
⋅기아 시와 같이 체내 포도당 농도가 낮아졌을 경우, 당뇨병이 심해 혈중 포도당 농도는 높으나
조직으로 포도당이 들어가지 못할 경우 → 지방산의 산화로 생성된 아세틸 CoA의 농도가 높아
져서 일어남
⋅아세틸 CoA 농도 증가 → 피루브산 탈수소효소 키나아제 활성화 → 피루브산 탈수소효소의 인
산화 활성 저하, 즉 피루브산이 아세틸 CoA로 전환되는 정도가 감소함
⋅아세틸 CoA에 의해 피루브산 카르복실화효소 활성 증가 → 피루브산이 옥살아세트산으로 전환
되어 당신생에 이용할 수 있음
⋅결과적으로 지방산이 산화 → 아세틸 CoA 농도 증가 → 케톤체 생성 → 뇌조직과 같이 포도당
을 필요로 하는 조직에서 케톤체를 에너지원으로 이용, 간에서는 당신생이 증가하여 체내 포도
당을 유지하는데 도움을 줌
⋅두 개의 아세틸 CoA가 티올라제의 작용에 의해 결합 → 아세토아세틸 CoA가 생성
→ 또 한 개의 아세틸 CoA와 결합 → β-하이드록시-β-메틸 글루타릴 CoA(HMG CoA)를 생성
⋅이 과정에서 HMG CoA 합성효소가 작용함
2) 케톤체의 이용 ⋅케톤체 중에서 아세토아세트산과 β-하이드록시부티르산 → 혈액을 통해 근육, 심장, 뇌 및 신
장 등 다른 조직으로 이동 → 미토콘드리아 내에서 아세틸 CoA를 생성하여 에너지원을 제공함
⋅간에서는 β-케토아실 CoA 전이효소가 없어 케톤체를 이용하지 못함
4. 지방산의 생합성
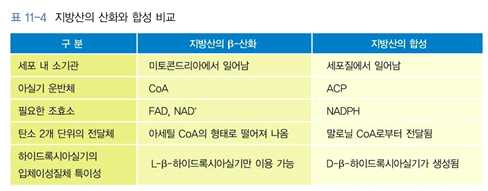
⋅체내에서 이용되는 지방산의 상당량은 식사를 통해 섭취하는 지방으로부터 공급
⋅체내에서 합성되지 못해 식사를 통해 섭취해야 하는 필수지방산
→ 리놀레산(linoleic acid), α-리놀렌산(α -linolenic acid)를 제외하고 대부분의 지방산들이
체내에서 합성될 수 있음
⋅지방산의 생합성은 아세틸 CoA에 탄소 2개 단위씩 결합하여 이루어짐
⋅지방산의 길이를 늘려 나가는 과정은 말로닐 CoA의 형태로 탄소 2개가 전달되고 이산화탄소가
떨어져 나가는 반응에 의해 일어남
⋅합성은 간, 지방조직, 유선에서 일어나며, 세포 내의 세포질에서 이루어짐
1) 지방산의 생합성 과정
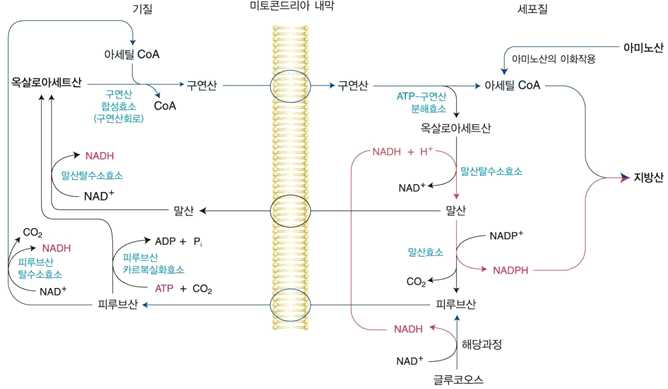
(1) 미토콘드리아에서 세포질로 아세틸 CoA의 이동
⋅포도당의 산화, 지방산의 산화, 케톤체의 대사 및 일부 아미노산의 대사과정에서 아세틸 CoA가
생성됨
⋅아세틸 CoA는 CoA부분 때문에 미토콘드리아막을 통과 불가
⋅잉여의 아세틸 CoA가 세포질로 이동하여 지방산 합성에 이용되어야 할 경우, 아세틸 CoA가
옥살아세트산과 결합하여 구연산의 형태로 전환되어 미토콘드리아 밖으로 나오게 됨
⋅구연산이 미토콘드리아 밖으로 나오는 것은 미토콘드리아 내의 구연산 농도가 높을 때
⋅ATP가 충분핛 경우 α–케토글루타르산 생성 저해 구연산 축적
(2) 아세틸 CoA 카르복실화효소의 작용에 의한 아세틸 CoA로부터 말로닐 CoA의 생성

⋅두 단계의 과정으로 일어남
1단계 : 카르복실화효소의 리신 잔기에 결합되어 있는 조효소인 비오틴이 카르복실화
2단계 : 아세틸 CoA가 비오틴에 결합되어 있는 카르복실기를 전달받아 말로닐 CoA를 생성
⋅지방산의 생합성에서 중요핚 조절 반응으로서 여러 가지 방법에 의해 조절됨
(3) 지방산 합성효소의 작용

⋅지방산 합성효소(FAS) 작용에 의해 팔미트산이 생성되는 과정은 7단계의 효소 반응에 의해 일
어남
⋅지방산 생합성 사이클에서 각 1분자의 H2O가 생성
⋅7회의 사이클 후 팔미토일-ACP로부터 팔미트산이 떨어져 나올 때 H2O가 사용되기 때문
→ H2O는 6분자 생성
① 아세틸 CoA의 아세틸기가 지방산 합성효소의 아실운반단백질(ACP)에 결합
② ACP에 결합되어 있던 아세틸기가 β-케토아실 합성효소의 시스틴 잔기에 있는 -SH기로 이동
③ 말로닐 CoA가 ACP에 결합
④ 말로닐 CoA에서 CO2가 떨어져 나옴 → 이 때, ACC의 작용에 의해 아세틸 CoA에 결합되어져 있던 것
⑤ ACP에 결합되어 있는 케토아실기가 β-케토아실 환원효소의 작용에 의해 환원(NADPH)되어
D-β-하이드록시부티릴-ACP가 됨
⑥ β-하이드록시아실-ACP 탈수소효소의 작용에 의해 물 분자가 빠져나오면서 탄소 2번과 3번
사이에 이중결합이 생겨 α, β-트랜스-부테노일-ACP가 생성됨(에노일)
⑦ 에노일-ACP 환원효소의 작용에 의해 환원(NADPH)되어 아실기로 전환 → 부티릴-ACP가 생성
⑧ 탄소 4개의 아실기가 –케토아실 합성효소의 시스틴 잒기에 있는 –SH기로 이동하고 ACP에
말로닐 CoA가 다시 와서 결합하면서 다시 위와 같은 반응을 반복하여 탄소 6개의 아실기를
생성함
⑨ 7회의 사이클을 거치면 탄소 16개의 팔미토일-ACP가 생성, 팔미토일 티오에스터라아제의 작
용에 의해 팔 미토일기가 ACP로부터 떨어져 나와 팔미트산이 생성됨. 이 때, 한 분자의 H2O
가 필요함

⋅아세틸 CoA로부터 말로닐 CoA가 생성되고, 아세틸 CoA와 말로닐 CoA로부터 지방산이 생성
되는 전체 반 응식을 팔미트산을 예로 들면 다음과 같은 반응식이 일어남
2) 지방산 생합성의 조절 (1) 아세틸 CoA 카르복실화효소(ACC)의 조절
⋅ACC는 지방산의 생합성에서 속도조절단계에 작용하는 효소
⋅다른자리입체성(알로스테릭)조절
- 구연산은 ACC의 활성증가
- 효소 활성 최대 속도를 증가시킴
- 효소 활성 억제제의 결합을 막고, 중합체가 형성 되도록 함
⋅미토콘드리아 내의 아세틸 CoA의 농도 증가 → 세포질 내의 구연산의 농도 증가
→ 지방산 생합성 증가
⋅지방산 합성 과정의 산물인 긴 사슬 아실 CoA는 ACC의 활성을 억제함
⋅인산화에 의한 조절
- ACC는 인산화가 되면 불활성화가 되고, 탈인산화가 되면 활성형이 됨 → 호르몬에 의해 조절
- ACC를 인산화 시키는 호르몬 : 글루카곤, 에피네프린, AMP에 의해 활성화되는 키나아제(AMPK)
- 세포 내에너지가 낮아짐 → AMPK 활성화 → ACC 인산화 → ACC 활성감소 → 지방산 합
성 감소
(2) 지방산 생합성에 관여하는 효소의 발현 조절
⋅ACC와 FAS의 발현은 호르몬, 열량 섭취에 의해 조절될 수 있음
⋅고당질식을 장기간 섭취 → 지방산 생합성에 관여하는 효소의 양이 간에서 증가
⋅열량이 적은 식이섭취, 기아 시, 다가 불포화지방산 섭취 시 → 효소들의 양 감소
3) 지방산의 연장과
불포화지방산의 합성 (1) 지방산의 연장
⋅세포질에서 지방산 합성효소(FAS)에 의한 반응에서 생성되는 것은 탄소 16개의 팔미트산임
⋅탄소 16개보다 더 긴 지방산
→ 팔미트산에 탄소 2개씩을 덧붙이는 반응에
→ 활면소포체, 미 토콘드리아에서 장쇄화효소의 작용에 의해 일어남
⋅활면소포체 : 탄소 2개가 말로닐 CoA와의 탈탄산반응에 의해 공급
→ 지방산 생성효소의 반응과 유사
⋅미토콘드리아 : 직접 아세틸기가 더해짐 → 지방산의 β-산화의 반대과정과 비슷
(2) 불포화지방산의 합성
⋅불포화지방산의 생성은 소포체의 막에서 탈포화효소의 작용에 의해 포화지방산에 이중결합이
만들어져서 일어남
⋅Δ9-, Δ6-, Δ5-, Δ4-지방산 아실 CoA 탈포화효소가 있으며, C9 위치 이 후에는 이중결합을
만들 수가 없음
⋅올레산(18:1 .9)의 합성은 가능, 리놀레산(18:2 .9,12 )의 합성은 불가능
⋅식사로부터 섭취한 리놀레산과 α-리놀렌산은 장쇄화효소와 탈포화효소의 작용에 의해
⋅리놀레산 → γ-리놀렌산과 아라키돈산으로 전환될 수 있음
⋅α-리놀렌산 → EPA와 DHA로 전환될 수 있음
5. 복합지질의 대사
⋅복합지질은 골격구조에 지방산이 공유결합된 것
⋅글리세롤지질(글리세롤 골격구조), 스핑고지질(스핑고신 골격구조)
⋅생물체에 의해 합성 or 섭취한 대부분의 지방산은 에너지 저장을 위한 중성지방으로 되거나
세포막 인지질 구성성분으로 결합됨
⋅생물체의 빠른 성장 동안 새로운 막의 합성 → 막인지질의 생성을 필요로 함
⋅활발하게 성장 X, 지방산 공급이 많을 때 → 대부분 지방으로 저장
⋅두 경로(글리세롤인지질, 트리아실글리세롤)는 공통의 중간대사물, 즉 글리세롤의 지방 아실 에
스테르의 생성으로부터 시작함
1) 글리세롤지질의 생합성 ⋅해당과정의 대사산물인 다이하이드록시아세톤인산(DHAP)로부터
→ 글리세롤 3-인산이 생성됨
(1) 중성지방(트리아실글리세롤, triacylglycerol)의 생합성
⋅다이하이드록시아세톤인산(DHAP) 또는 글리세롤로부터 글리세롤 3-인산이 생성됨
⋅글리세롤지질 생합성의 첫 번째 단계 : 글리세롤 3-인산의 2개의하이드록시기 + 2분자의 지방산 아
실 CoA
→ 다이아실글리세롤 3-인산(포스파티드산)을 생성함
⋅포스파티드산은 세포에 소량만 존재하지만 지질 생합성에서 중요한 중간물질로서 중성지질이나 글리
세롤인지질로 전환될 수 있음
⋅포스파디트산은 세포에 소량만 존재하지만 지질 생합성에서 중요한 중간물질로서 중성지질이나
글리세롤인 지질로 전환될 수 있음
⋅포스파티드산 → 다이아실글리세롤이나 CDP-다이아실글 리세롤로 전환 → 이로부터 모든 글리
세롤인지질 합성
⋅중성지방 → 주로 지방조직, 간 및 소장에서 합성되고 주요 에너지 저장물질로서 작용함
⋅식사로부터 온 중성지방 : 리파아제에 의해 → 2-모노아실 글리세롤로 분해 → 아실화반응을
촉매하는 아실전이효소에 의해 → 새로운 중성지방이 합성됨
| ※ 탈포화효소 : 비헴철(non-heme iron)을 가지고 있는 막효소, 전자전달계 시스템의 일부 |
(2) 글리세롤인지질(glycerophospholipid)의 대사
① 글리세롤인지질의 생합성
⋅포스포파티딜에탄올아민(PE)의 생합성 :
에탄올아민의 인산화 포스포에탄올아민 형성 → CTP로부터 시티딜기 이동에 의해
→ CDP-에탄올아민, 피로인산(PP) 생성 → 포스포에탄올아민 전이효소에 의해
→ 포스포 에탄올아민을 다이아실글리세롤 골격에 연결
⋅포스파티딜세린(PS)은 포스파티딜에탄올아민의 에탄올아민 세린으로 교환하는 반응에
의해 생성됨
⋅포스파티딜콜린(PC)의 생합성 : 포스파티딜에탄올아민 생합성과정과 유사함
⋅이 경로에서 필요한 모든 콜린은 식사로부터 얻어져야 함
2) 글리세롤 인지질의 분해
⋅모든 조직과 체액에서 발견되는 포스포리파아제에 의해 글루세롤인지질의 포스포다이에스터 결
합을 가사분 해하여 이루어짐
⋅포스포리파아제의 종류에 따라 인지질 내에서 가수분해되는 부위가 다름
⋅글리세롤인지질의 1번 또는 2번 탄소로부터 지방산이 떨어져 나가면 리소포스포글리
세리드 생성(리소포스포 리파아제의 작용을 받게 됨)
⋅포스포리파아제의 작용에 의해 전령으로 작용하는 물질(다이아실글리세롤, 이노시톨
1,4,5-삼인삼) 또는 전 령의 전구체 물질(아라키돈산, arachoidonic acid)이 방출됨
3) 스핑고지질의 대사
(1) 스핑고지질의 생합성
⋅스핑고지질은 신경조직에 고농도로 존재
⋅세라마이드(ceramide)는 모든 다른 스핑고지질의 기본 골격
(예. 스핑고미엘릮: 포스포콜린 + 세라미드)
(2) 스핑고지질의 분해
⋅리소좀 효소인 스핑고미엘린 가수분해 → 포스포릴콜린을 제거 → 세라미드로 변함
⋅세라미드 + 세라미드 분해효소 → 스핑고신 + 유리지방산으로 분해
⋅세라미드 : 스트레스에 대한 반응에 관여
⋅스핑고신 : 단백질 키나아제 C의 활성을 억제함
<개념확인문제>
문제 1. 다음 화합물 중 케톤체(ketone body)에 속하는 물질은? [2016년_27번]
가. β-하이드록시부티르산 나.아세톤 다.아세토아세트산 라.옥살로아세트산
① 가, 나, 다, 라
② 가, 나, 다
③ 가, 다, 라
정답 ② ④ 나, 다, 라
문제 2. 다음 화합물 중 케톤체(ketone body)에 대한 설명으로 옳은 것은? [2017년_28번]
① 하이드록시부티르산, 아세토아세트산, 아세톤 등이 있다.
② 아세톤은 혈액을 통해 다른 조직으로 이동하여 에너지를 생성한다.
③ 케톤체의 생성은 신장에서 이루어진다.
정답 ① ④ 보통 체내 포도당이 충분할 경우에 다량 생성된다.
문제 3. 스테아르산은 탄소수가 18개인 포화지방산이다. 이 스테아르산이 완전히 산화될 때 몇 회의 β-산
화과정과 몇 개의 아세틸 CoA가 구연산회로를 통과하며, 결국 최종 생성되는 ATP의 개수는
몇 개인지 순서대로 표시한 것은? [2016년_2번]
① 9 - 8 - 120
② 8 - 9 - 120
③ 9 - 8 - 150
정답 ② ④ 8 - 9 – 150
문제 4. 중성지방을 완전히 가수분해하면 1 분자의 글리세를과 ( ) 분자의 지방산을 얻는다. 다음 중 괄
호에 들어갈 숫자로 옳은 것은? [2017년_8번]
정답 ① ① 3 ② 4 ③ 5 ④ 6
문제 5. 중성지방을 완전히 가수분해하면 ( A ) 분자의 글리세롤과 ( B ) 분자의 지방산을 얻을 수 있다.
A와 B를 더하면? [2018년_25번]
정답 ② ① 3 ② 4 ③ 5 ④ 6
문제 6. 콜레스테롤 합성의 중간 물질로만 나열한 것은 무엇인가? [2016년_6번]
① HMG CoA, 메발론산, 스쿠알렌
② 아세틸 CoA, 피루브산, 스쿠알렌
③ HMG CoA, 메발론산, 세라미드
정답 ① ④ 아세틸 CoA, 피루브산, 세라미드
문제 7. 지방산 산화의 첫 단계인 지방산의 활성화는 미토콘드리아 밖에서 일어난다. 이후 더욱 산화되기 위해
서는 미토콘드리아 안으로 들어가야 하는데, 이 때 필요한 운반체(carrier)로 옳은 것은?[2018년_14번]
정답 ② ① 오르니틴 ② 카르니틴 ③ 시트룰린 ④ 크레아티닌
문제 8. 다음은 A라는 물질이 합성될 경우에 나타나는 그 중간 물질들을 나열한 것이다. A라는 물질은
다음 중 무엇인가? [2017년_6번]
HMG CoA, 메발론산, 스쿠알렌
① 글리코겐 ② 헤모글로빈
정답 ③ ③ 콜레스데롤 ④ 비타민 C
문제 9. 지방산의 연장, 장쇄화 과정에 대한 설명으로 옳은 것은?
① 세포질에서 지방산 합성효소에 의해 생성된 스테아르산으로부터 시작한다.
② 미토콘드리아에서는 아세틸기가 직접 더해져 지방산의 베타산화의 반대과정과 유사하다.
③ 활면소포체에서는 탄소가 3개씩 더해진다.
정답 ② ④ 미토콘드리아와 활면소포체에서 일어나는 연장과정은 동일하다.
문제 10. 다음 중 지방산의 생합성에 대한 설명으로 옳은 것은?
㉮ 지방산 생합성을 통해 8~12개의 사슬길이를 가진다.
㉯ 말로닐 CoA를 통해 지방산 사슬이 2탄소 단위씩 연속적으로 증가하는 과 정이다.
㉰ 지방산 환원반응의 환원제로 NADH가 사용된다.
㉱ 아세틸 CoA의 카르복실화 반응으로 말로닐 CoA가 생성된다.
정답 ③ ① ㉮㉯㉰ ② ㉮㉰ ③ ㉯㉱ ④ ㉮㉯㉰㉱
문제11. 지방산 산화의 첫 단계는 미토콘드리아 밖에서 일어나며, 더 산화되기 위하여 미토콘드리아 안으
로 들어가야 한다. 이런 이동은 특별한 운반계에 의해 일어나는데, 이 이동에 필요한 물질로 옳은
것은?
정답 ③ ① 크레아틴 ② 오르니틴 ③ 카르니틴 ④ 담즙산
문제12. 탄소수가 18개인 스테아르산 지방산이 β-산화과정을 통해 최종생산물 아세틸-CoA로 될 때, 몇
개의 아세틸-CoA가 생성되는가?
정답 ④ ① 1개 ② 3개 ③ 5개 ④ 9개
문제13. 다음 중 지단백질 밀도의 수치와 동맥경화와 관련있는 것으로 옳은 것은?
㉮ HDL의 상승 ㉯ HDL의 저하 ㉰ LDL의 하강 ㉱ VLDL의 상승
정답 ③ ① ㉮㉯㉰ ② ㉮㉰ ③ ㉯㉱ ④ ㉮㉯㉰㉱
※ 부족하지만 글의 내용이 도움이 조금이라도 되셨다면, 단 1초만 부탁드려도 될까요? 로그인이 필요없는 하트♥(공감) 눌러서 블로그 운영에 힘을 부탁드립니다. 그럼 오늘도 행복한 하루 되십시오^^
'생화학' 카테고리의 다른 글
| 식품, 영양 관련 쉽게 정리한 생화학 핵심 요점 요약 정리 13. 아미노산 및 질소대사(1) (0) | 2022.05.12 |
|---|---|
| 식품, 영양 관련 쉽게 정리한 생화학 핵심 요점 요약 정리 12. 지질 대사(2) & 아미노산 및 질소대사(1) (0) | 2022.05.11 |
| 식품, 영양 관련 쉽게 정리한 생화학 핵심 요점 요약 정리 10. 지질 (0) | 2022.05.09 |
| 식품, 영양 관련 쉽게 정리한 생화학 핵심 요점 요약 정리 9. 전자전달계와 산화적 인산화 (0) | 2022.05.08 |
| 식품, 영양 관련 쉽게 정리한 생화학 핵심 요점 요약 정리 8. 구연산회로 (0) | 2022.05.07 |