제 11 장. 지방질 대사
| ⦁acetate mevalonate pathway : acetyl CoA를 원료로 하여 메발론산(mevalonate)을 거쳐 스테로이드가 합성되는 경로이다. ⦁fatty acid synthase(FAS) 복합체계 : 지방산 합성에 필요한 효소들의 복합체 ⦁methyl erythiol phosphate pathway : pyruvate를 원료로 하여 methyl erythriol을 거쳐 isoprenoid가 합성되는 경로로서 isoprenoid는 식물의 생리활성물질 합성에 이용된다. ⦁α-산화(α-oxidation) : 지방산의 α 탄소가 산화되어 이산화탄소로 소실되는 반응 ⦁β-산화(β-oxidation) : 지방산이 산화되어 acetyl CoA를 생산하는 반응인데 지방의 β 탄소, 즉 카르복시기에서 두 번째 탄소가 산화된다. ⦁ω-산화(ω-oxidation) : 지방산의 ω 탄소가 산화하여 discarboxylic acid가 형성되는 반응 ⦁고밀도 리포단백질(HDL) : 혈액에서 지방질을 운반하는 지질단백질로서 콜레스테롤과 중성지방 함량이 낮으며, 콜레스테롤을 조직,세포로부터 간으로 운반한다. ⦁아실기 운반단백질(acly carrier protein: ACP) : fatty acid synthase(FAS) 복합체계의 일부로서 합성과정 중에 있는 acyl기를 고정한다. ⦁유미입자(chylomicron) : 동물체 소장의 점막상피세포로 흡수된 지방산과 mono-glyceride가 triglyceride로 합성된 후 지질단백질,콜레스테롤과 혼합하여 형성한 크기 1㎛이하의 작은 입자로 임파관으로 흡수되어 체내로 이동한다. ⦁저밀도 리포단백질(LDL) : 혈액에서 지방질을 운반하는 지질단백질로서 콜레스테롤 함량이 높으며, 콜레스테롤을 간에서 조직, 세포로 운반한다. ⦁케톤증(ketosis) : 케톤체가 너무 많이 생산되어 혈액 중의 아세토아세트산과 β-히드록시부티르산의 농도가 높아져 체액이 산성화함으로써 나타나는 현상이다. ⦁케톤체(ketone body) : 케톤기(-C=O)를 가지는 물질을 의미한다. |
11.1. 지방질 대사골격
○ 지방질(lipid)
- 구성분자에 반드시 지방산을 함유한 유기화합물
- 탄수화물, 단백질과 생물체의 주요 성분
- 물에 용해되지 않고, 유기용매에 용해됨
- 에너지 값(약 9kcal/g)이 다른 영양소에 2배 이상
- 단순지방질, 복합지방질 및 유도지방질로 구분
○ 지방질 대사
- 중성지방(triacylglycerol) : 지방조직 90% 차지
- 인지질(phospholipid) : 단백질과 함께 생체막 구성
- 스테롤(sterol) : 생체막 구성 및 스테로이드생합성 전구체
- 당지질(glycolipid) : 세포벽 및 항원성물질(혈액형 결정) 성분
1) 동물

㉮ 동물이 섭취한 지방질은 소화기관에서 가수분해 후 흡수되어
이용되고, 세포의 지방질은 호르몬에 의해 활성화된 지방질
가수분해효소에 의해 분해된 후에 이용된다.
㉯ 동물체에서는 탄수화물 에너지원이 소모된 후에 지방질이 에
너지원으로 이용된다. 탄수화물에너지원이 과다할 때에는 지방
질로 전환되어 저장된다.
㉰ 지방질 분해물은 물질 합성원료로 사용될 수 있다.
2) 식물
식물의 경우 지방질은 주로 물질합성의 원료로 사용되며, 유량종자에서는 지방질이 에너지원과 물질 합성원료로 사용된다.
11.2. 지방질의 분해
○ 지방의 소화·흡수
- 위 : 리파아제 → 중성지방의 10∼30% 정도가 소화
- 췌장 : 리파아제, 코리파아제, 콜레스테롤 에스터라아제, 포스포리파아제
- 십이지장 : 작은 기름 방울 형태 지방 → 유화액 형성
· 유화액은 표면적은 증가시켜 췌장 분비 소화효소의 촉진
1) 중성지방
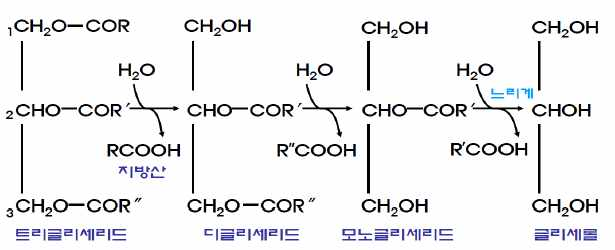
중성지방의 가수분해 트 리글리세리드 → 글리세롤 + 지방산 췌 장 분비 소화효소의 촉진
(1) 동물
① 식이지방
담즙산염에 의해 유화된 후 소화액으로 분비된 지방 분해효소에 의해 지방산과 글리세롤로 분해된다.
② 지방세포 지방
아드레날린, 글루카곤, 부신피질 자극호르몬 등 호르몬 자극에 의해 지방 분해효소가 인산화되어 활성화한 후 지방세포의 지방을 분해한다.
(2) 식물
종자가 발아할 때 지방 분해효소가 지방을 분해한다.
2) 인지질
포스포리파아제 A, B, C, D에 의해 지방산, 제2알코올, 인산
부위가 모두 가수분해된다.
3.동물의 식이지방질의 흡수, 운반
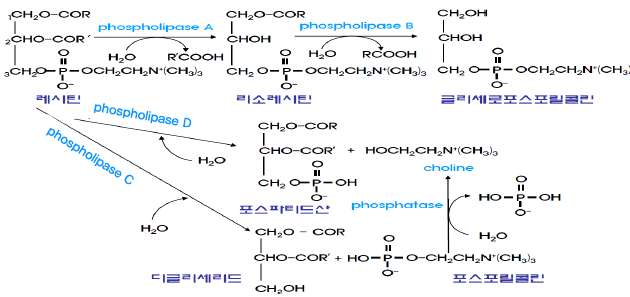
인지질의 가수분해 (포스포리파아제 A, B, C, D에 의해 가수분해)
㉮ 분해된 지방질 물질들은 소장 상피의 점막세포에서 흡수된 후
중성지방으로 형성되어 지질을 혈액에서 운반하는 단백질인 아포리포단
백질(apolipoprotein)과 결합하여 유미입자를 형성하여 임파관으로 흡
수된 후 흉선을 거쳐 혈관으로 이행하여 세포에 도달한다. 조직의 세포
에 도달하면 리포단백질 지방질 가수분해효소에 의해 다시 분해된 후
세포로 들어간다. 저급지방산은 직접 흡수되어 임파관을 거치지 않고
혈관으로 흡수된다.
㉯ 아포리포단백질은 식이지방질을 조직과 세포로 운반하는 기능 외에 과잉
으로 섭취한 에너지원을 간장으로부터 조직, 세포로 운반하는 기능도
가지고 있다. 과잉으로 섭취한 에너지원은 간에서 중성지방으로 전환되는
데 아포리포단백질이 중성지방과 결합하여 혈액을 따라 조직, 세포로 운반한
다. 아포리포단백질은 중성지방 외에 콜레스테롤도 결합하여 운반하는데 이를
초저밀도 리포단백질이라 한다. 초저밀도 리포단백질이 근육과 지방조직에 도
달하면 중성지방이 분해, 분리되어 이용되며, 저밀도 리포단백질로 형성되는데
이는 콜레스테롤 함량이 높다.
㉰ 초저밀도 리포단백질에서 중성지방이 제거되면 콜레스테롤 함량이 높은 저밀도 리포단백질이 형성되는데
이는 콜레스테롤을 간으로부터 조직, 세포로 운반한다. 혈관에 축적하여 심혈관계 질환의 원인이 된다.
㉱ 콜레스테롤과 중성지방이 많이 제거되어 함량이 낮은 아포리포단백질은 고밀도 리포단백질이며 조직, 세포
에서 콜레스테롤을 간으로 운반한다. 간으로 운반된 콜레스테롤은 담즙산 합성원료로 사용되거나 분해된
다.
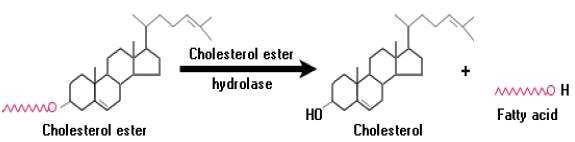
11.2.3 콜레스테롤 에스테르 (콜레스테롤 에스터라아제 → 콜레스테롤 + 지방산)
11.3. 지방분해물의 이용
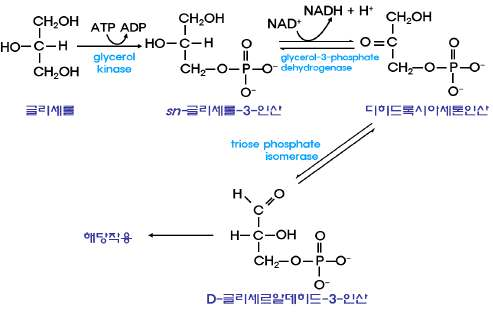
11.3.1 글리세롤
㉮ 글리세롤-3-P로 전환되어 중성지방 합성에 이용된다. ㉯ 글리세르알데히드-3-P로 전환되어 에너
지원으로 이용된다.
11.3.2 지방산
11,3.2.(1) 동물
1) 지방산 산화 (fatty acid oxidation) - 동물
- 지방산은 β산화 → 아세틸CoA 전환 → 물질 합성 & 에너지
- 반응 장소 : 미토콘드리아 내막 안쪽의 기질
- 반응 경로
· 시토졸의 지방산 → 지방산 CoA 형성 → 미토콘드리아 이동 → 미토콘드리아 기질에서 β산화 → 에너지 생산
지방의 가수분해 (트리글리세리드 → 글리세롤 + 지방산)
㉮ 지방산 CoA의 형태로 미토콘드리아 기질로 이동한 후
지방산의 β-탄소가 산화하는 β-산화과정을 통하여 산화
되어 acetyl CoA로 전환된 후 물질 합성원료로 사용되
거나 에너지원으로 이용된다.

지방산의 미토콘드리아 기질로 이동
㉯ β-산화 1회전에서 탄소 수가 2개인 acetyl CoA,
FADH2, NADH 각 한 분자씩 생산된다. 그러므로 2n개
의 짝수탄소를 가지는 포화지방산은 (2n/2)-1회의 β-산
화를 거쳐 NADH와 FADH2를 각기 n-1개씩 생산하고,
acetyl CoA는 n개 생산한다. acetyl CoA는 크렙스회로
에서, FADH2와 NADH는 전자전달계에서 ATP를 생산
한다.
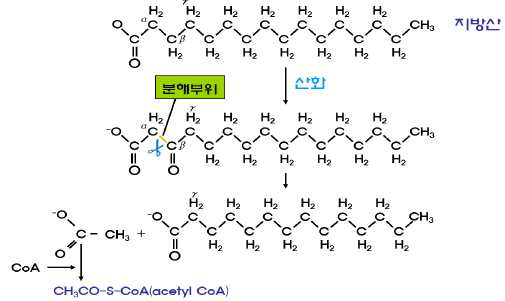
지방산의 α-탄소와 β-산소 사이의 분해
㉰ 홀수개의 탄소로 구성된 지방산이 β-산화과정을 거치
면 마지막 지방산 CoA는 탄소 수가 3개인 propionyl
CoA인데 이는 이산화탄소와 반응하여 숙시닐 CoA로
전환된 후 크렙스회로에서 이용된다.
㉱ 불포화지방산은 불포화결합 부위를 β-산화과정의 중간물질인
enoyl-CoA 와 동일한 형태로 바꾸는 것이 필요하다. oleic
acid의 경우에 세 번의 β-산화 후 isomerase에 의해
enoyl-CoA와 동일한 구조를 가지게 됨으로써 β-산화에 의해
분해될 수 있다.
㉲ α 탄소, ω탄소가 산화하는 경우도 있다.
2) β-산화
2.1 포화지방산 β-산화
- 지방산을 에너지원으로 사용하기 위한 아세틸CoA
생성 대사(지방산의 에너지 생산 대사과정)
○ 지방산 β산화 - 4가지 주요 반응
① 아실CoA 탈수소효소 : FAD가 FADH2로 환원
② 에노일CoA 수화효소 : 수화(이중결합의 β위치에 -OH 첨가)
③ 하이드록시아실CoA 탈수소효소 : NAD가 NADH로 환원
④ 티올라아제 : 아세틸CoA 와 탄소 2개가 짧아진 지방산CoA
→ 글루코오스와 스테아르산의 ATP생산효율을 비교하면 탄소 하나
당 ATP를 5.3분자 생산하고, 분자량으로 보면 0.18분자 생산
된다. 스테아르산은 탄소하나당 ATP를 6.7분자생산하고 분자
량으로 보면 0.42분자의 ATP를 생산한다. 탄소하나당 ATP의
생산량은 유사하지만 분자량당 ATP생산량은 지방산이 탄수화
물의 2배이상이다. 그러므로 지방이 탄수화물보다 에너지 저장
량이 대단히 많다.
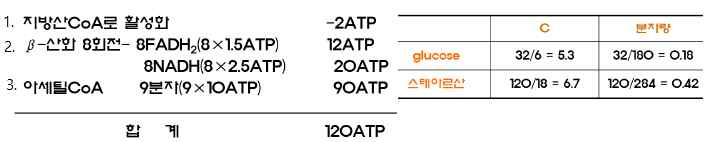
※ 스테아르산(18:0)이 산화결과 ATP 생성
○홀수탄소 지방산의 산화
- 자연계 존재 대부분의 지방산은 짝수 개의 탄소
- 일부 식물, 해양동물, 미생물은 홀수 탄소의 지방산 존재
- 산화 결과물 : (탄소수-3)/2개의 아세틸 CoA와 1개의 프로피오닐 CoA
- 프로피오닐 CoA→숙시닐 CoA전화→크렙스회로 이용

홀수탄소 지방산의 β-산화
2.2 불포화지방산의 β산화 : 불포화지방산은 이중결합이 에노일 CoA에서의 형
태로 전환되어 β-산화과정에서 이용된다
- 불포화지방산의 이중결합 : cis형
- β산화과정 2번째 물질인 에노일 CoA 수화효소 : trans형
- 에노일 CoA와 동일한 이중결합과 위치(C2∼C3) 전환
- 포화지방산 vs. 불포화지방산 산화 결과
· 이중결합으로 FADH2 생성과정이 생략됨
· 포화지방산의 산화에 비해 ATP가 덜 생성됨
11,3.2.(2) 식물
○ 지방산 산화 (fatty acid oxidation) - 식물
- 반응 장소 : 잎의 퍼옥시좀, 발아 종자의 글리옥시솜
- 미토콘드리아에는 β산화체계가 없음
- FADH2가 산소와 반응하여 과산화수소 생성
- 지방산의 β산화 생성물인 아세틸 CoA는 물질 합성 원료
※ 식물에서의 지방산 β산화는 에너지생산 목적이 아닌 물질 합성원료를 공급하는 목적임
㉮ 식물의 지방산 β-산화는 잎의 퍼옥시좀(peroxisome)과 발아하는 종자의 글리옥시솜(glyoxysome)에서 일어나며, 미토콘드리아에는 β-산화체계가 없다.
㉯ 식물에서의 지방산 β-산화과정은 동물의 미토콘드
리아에서 일어나는 경우와 거의 유사하나 생성된
FADH2가 에너지원으로 사용되지 않고 산소와 반응
하여 FAD를 재생하고, NADH는 퍼옥시좀 밖으로 나
가 미토콘드리아의 전자전달계에서 산화되어 ATP를
생산한다.
㉰ 글리옥시솜과 퍼옥시좀에는 크렙스회로 대사체계가
존재하지 않기 때문에 지방산의 β-산화에 의해 생성
된 acetyl CoA는 물질 합성의 원료로 사용된다. 그
러므로 식물에서의 지방산 β-산화는 에너지생산에 목
적이 있지 않고 물질 합성원료 공급에 목적이 있다.
그러나 일부 acetyl CoA는 미토콘드리아로 이동하여
에너지원으로 사용될 수도 있다.

지방산 β-산화
11,3.2.(3) 케톤체 형성
○ 케톤체(ketone body, ketone)
- (전환)아세토아세트산, β-하이드록시부트르산, 아세톤
- 생성 장소 : 간
- 혈중 케톤체 농도가 높으면 혈액 pH 낮아지고 체액산성화
- 아세틸 CoA가 크렙스회로에 이용될 수 있는 옥살로아세트산이
부족하여 케톤체가 형성됨
- 글루코오스 신생합성 증가에 따른 옥살로아세트산 소모
㉮ 지방산의 β-산화에 의해 생산된 acetyl CoA가 크렙스회로에서
이용될 수 있기 위해서는 옥살로아세테이트가 필요하다. 지방이
과다하게 에너지원으로 이용되면 옥살로아세테이트가 충분치 않
아(부족) acetyl CoA의 일부는 크렙스회로에 이용되지 못하고
아세토아세트산, 아세톤, β-히드록시부티르산 등으로 전환된다.
이들을 케톤체라 한다. β-히드록시부티르산은 케톤체가 아니지
만 아세토아세트산, 아세톤과 함께 케톤체라 한다.→ 에너지원
으로 사용되지 못하는 아세틸CoA가 많아 케톤증 발생
㉯ 케톤체들은 체액을 산성화하여 몸체에 심각한 문제를 일으키며,
이러한 현상을 케톤증이라 한다.(혈액의 산성화)

케톤체의 생성
11.4. 지방질 합성
1) 포화지방산
㉮ 동물에서는 지방산이 간, 지방세포, 수유 중인 유선세포의 시토
졸에서 합성된다. 식물은 시토졸에서 지방산이 합성되지 않고
엽록체의 스트로마에서 지방산이 합성된다. 이는 광합성의 명반
응에서 생산된 NADPH를 이용해야 하기 때문이다.
리놀렌산(α-linolenic acid)을 합성하지 못한다. 그러므로 이 두 지방산은 동물
에게 필수지방산이다. 식물은 이 두 지방산을 모두 합성할 수 있다.
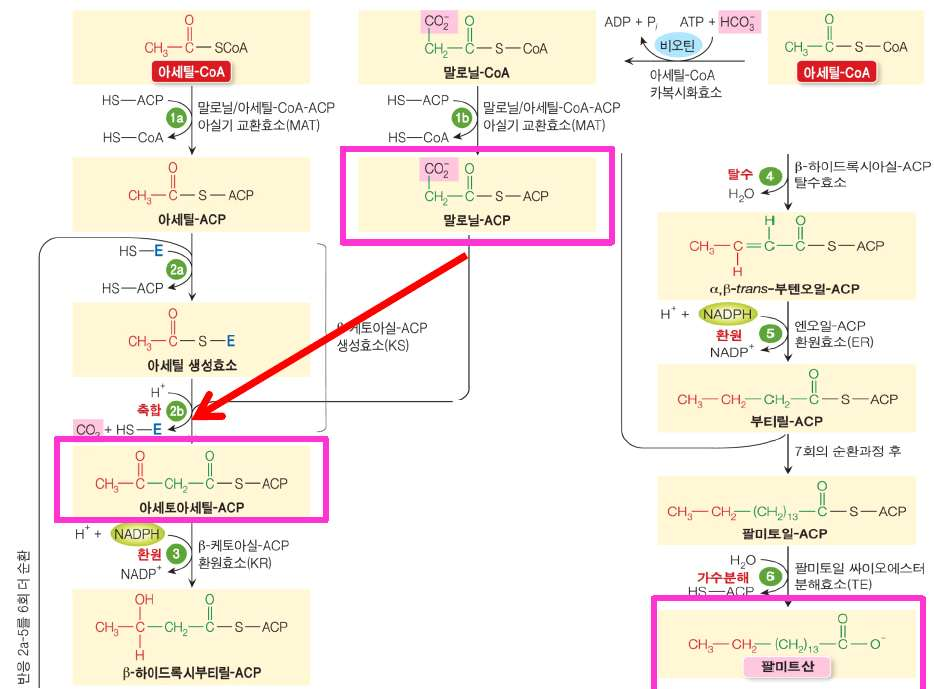
㉰ 동물에서는 불포화지방산 합성체계가 시토졸과 면하고 있는 소포체 표면에 존재하며, 식물에서는 엽록체, 소포체 내부에 존재한다㉯ acetyl CoA를 원료로 하여 여덟 가지의 효소로 구성된 지방산 합성기구(fatty
acid synthase: FAS)에서 NADPH에 의해 환원되면서 한 회전에 탄소가 2개
증가한 지방산이 합성된다.
※ 지방산은 acetyl-CoA를 원료로 하여 탄소가 첨가, 환원되면서 합성된다. 합성반
응은 환원되는 반응인데 NAD+와 FAD의 환원물이 NADH와 FADH2에 환원되지
않고 NADH에 의해 환원된다. 지방산 합성시 acetyl-CoA가 malonyl-CoA로
전환된 후 ACP(acyl carrier protein아실 운반 단백질)에 결합된다.
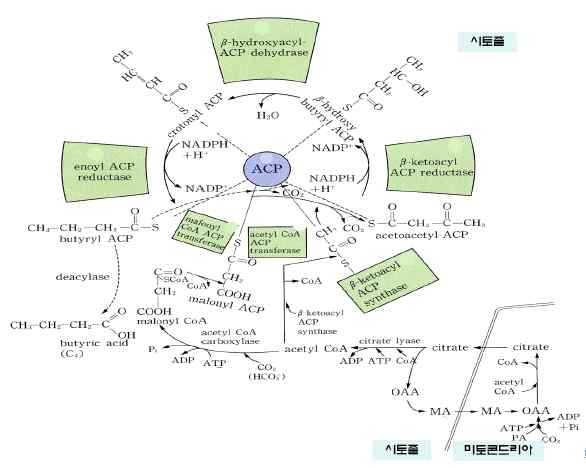
포화지방산의 합성
2) 불포화지방산
㉮ 불포화지방산은 포화지방산으로부터 수소가 제거되어 합성된다.
(팔미트산: 합성원료, ※합성경로)
㉯ 동물에서는 C9∼C10(Δ9)의 불포화결합은 형성할 수 있으나 C12∼C13(Δ12)와 C15∼
C16(Δ15) 위치의 불포화결합을 형성할 수 없기 때문에 리놀레산(linoleic acid)과
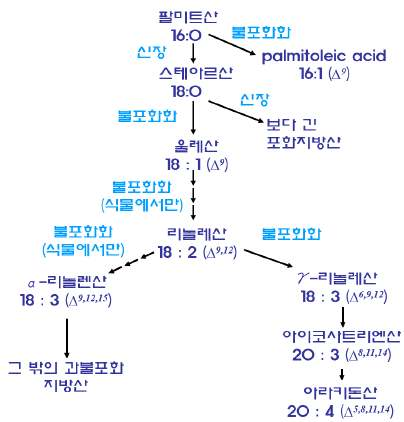
불포화지방산 합성경로
3) 중성지방 합성
㉮ 글리세롤 혹은 해당작용 중간생성물인 디히드록시아세톤인산으로부터 형성된 글리세롤-3-인산에 지방산이 C1, C2의 순서로 각기
두 분자의 ATP를 소모하면서 결합하고, C3의 인산이 제거된 후 지방산이 결합하여 중성지방이 합성된다.
㉯ 중성지방은 동물에서는 지방조직과 간 세포의 소포체 표면에서 합성되고, 식물에서는 소포체와 엽록체에서 합성된다.
㉰ 동물에서 혈중의 글루코오스 함량이 높으면 인슐린의 조절에 의해
acetyl CoA가 지방산으로 합성되면서 중성지방으로 전환되어 저
장된다. 인슐린이 부족하면 acetyl CoA가 지방산으로 전환되지
못하고 케톤체로 전환하면서 체중이 감소한다.

11.4.2. 인지질 합성
㉮ 인지질이 합성되는 방법은 두 가지이다. 중성지방 합성 중간생성물
인 포스파티드산에 CDP(cytidyne diphosphate, 시티딘이인산)가
결합하여 CDP-디글리세리드가 형성되고 이에 제2알코올류(원자
단)가 결합하면서 CMP가 분리되어 인지질(글리세로포스포지질)이
합성된다. 혹은 CDP가 제2알코올류(원자단)와 결합하고 디글리세
리드와 반응하여 인지질이 합성된다.
㉯ 인지질은 동물에서는 활면소포체 표면, 식물에서는 엽록체와 소포
체에서 합성된다. 즉 중성지방 합성장소와 동일하다.
㉰ 지방산이 중성지방을 합성할 것인가 인지질을 합성할 것인가는 생
물체의 필요에 따라 결정된다. 성장하는 생물체에서는 세포막성분
이 대량으로 필요하기 때문에 인지질 합성에 주로 이용되고, 성장을 완료한 생물체에서는 중성지방 합성에 이용된다.
1) 합성경로 및 장소
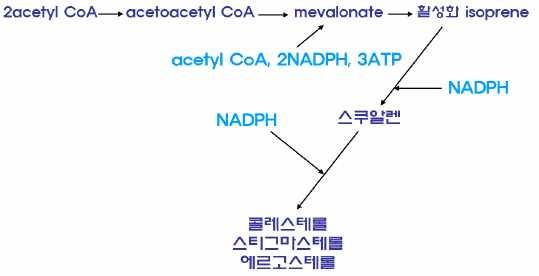
11.4.3 스테로이드 합성
(1) 동물
주로 간의 소포체에서 acetate mevalonate pathway에 의해
콜레스테롤이 합성된다.
(2) 식물
㉮ 소포체에서 acetate mevalonate pathway에 의해
stigmasterol이 합성된다.
㉯ 색소체(plastid) 시토졸에서는 methyl erythriol
phosphate(MEP) pathway에 의해 isoprenoid가 합성된다.
2) acetate mevalonate pathway
두 분자의 acetyl CoA가 축합하여 아세토아세틸 CoA가 합성되
고, 메발론산을 거쳐 동물에서는 콜레스테롤, 식물에서는 스티
그마스테롤, 진균류에서는 에르고스테롤이 합성된다.
3) methyl erythriol phosphate(MEP) pathway pyruvic acid를 원
료로 하여 methyl erythriol phosphate(MEP)을 거쳐
isopentenyl pyrophosphate을 합성한다. isopentenyl
pyrophosphate는 지베렐린, ABA(abscisic acid),
carotenoid 등 각종 생리활성 isoprenoid를 합성하는 원료이다.
<정리하기>
·동물체는 에너지원이 과다한 경우 지방질로 저장되며, 지방질은 에너지원
과 물질합성원료로 널리 이용된다. 식물은 필요한 만큼의 지방질만을 합성
하며, 지방질이 에너지원 혹은 다른 물질합성원료로 이용 되는 경우는 식
물의 종자가 발아 할 때를 제외하고는 없다.
·중성지방 분해물인 글리세롤은 글리세르알데히드-3-인산으로, 지방산은
β-산화에 의해 아세틸CoA로 전환되어 에너지원으로 이용된다. 지방산의 β
-산화에 의해 NADH, FADH2, 아세틸CoA가 생산된다.
·동물에서 지방산의 β-산화는 지방산으로부터 에너지를 생산하는 것을 목
적으로 하며, 식물에서는 물질합성원료로 사용하기 위함이다. 식물의 경우 종자에서는 β-산화에 의해 지방산으로부터 에너지를 생산한다.
·동물이 지방산을 에너지원으로 과다하게 사용하면 아세틸CoA가 과다하게 생산되어 케톤증을 유발한다.
·지방산은 아세틸CoA를 원료로 하여 합성되며, 포유동물은 지방산의 C12-C13과 C15-C16의 불포화결합을 형성하지 못하기 때문에
리놀레산과 리놀렌산이 필수지방산이다.
·스테로이드는 acetate mevalonate pathway에 의해 아세틸CoA가 축합하여 형성된 활성 이소프렌 단위를 원료로 하여 생성 된다.
식물은 methyl erythriol phosphate pathway에 의해 각종 생리활성 물질의 원료인 isoprenoid물질을 합성한다.
[교재 요약]
지방질 분해 산물 : 대부분 글리세롤과 지방산
지방산의 이용 양태
·동물 : 에너지원(주)과 물질 합성 원료(부)
·식물 : 주로 물질 합성 원료(종자 발아 제외)
지방질의 분해
① 동물
·식이지방 : 담즙산염(리파아제)에 의해 유화 가수분해
·세포지방 : 지방질 분해 효소가 호르몬에 의해 활성
② 식물 : 종자가 발아할 때 지방분해효소에 의해
글리세롤 :
①글리세롤-3-인산으로 전환하여 지방질 합성
②D-글리세르알데히드-3-인산으로 전환되어 해당작용 중간물질로서
에너지원으로 사용
지방산
·동물
①미토콘드리아 기질
②β-산화에 의해 아세틸CoA로 전환
③β-산화 1회전 ⇒
1아세틸CoA + 1탄소2개 감소한 지방산CoA + 1FADH2 + 1NADH
④짝수개 탄소 : (탄소수/2-1)회 β-산화, (탄소수/2) 아세틸CoA,
(탄소수), (FADH2 + NADH)
⑤탄소 1개당 ATP생산량은 탄수화물과 비슷하지만 분자량당 ATP
생산량은 2배이상
⑥혹수 개 탄소 : 프로피오닐CoA→숙시닐CoA→TCA회로
⑦불포화지방산은 에노일CoA의 형태로 전환되어야 β-산화과정에 이용함, 이성질화효소와 환원효소가 필요함.
·식물
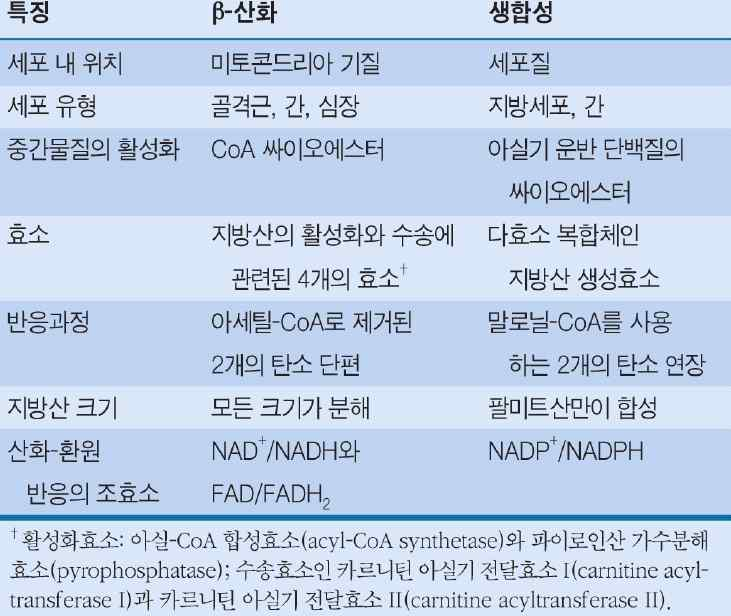
지방산 분해와 합성의 차이
②생성된 FADH2는 에너지원으로 이용되지 않고 산소
와 반응 하여 FAD를 재생산
③생성된 NADH는 퍼옥시좀을 나가 미토콘드리아의 전
자전달계에서 산화하여 ATP 생산
④퍼옥시좀과 글리옥시솜은 TCA회로가 없음→아세틸
CoA는 물질 합성의 원료로 사용
케톤체
①동물의 간에서 합성. 에너지원
②에너지원 과다→아세틸CoA과다→옥살로아세트산 부족→
남은 아세틸CoA가 케톤체로 전환→혈중 pH 감소
포화지방산 합성
①동물 : 유선세포의 시토졸
②식물 : 엽록체의 스트로마
③아세틸CoA를 원료로 탄소가 첨가, 환원되면는 합성
④탄소수가 16개인 팔미토일ACP(말미트산)일 때 종료
불포화지방산 합성
①동물 : 시토졸과 면하고 있는 소포체 표면
②식물 ; 엽록체와 소포체 표면
③포화지방산으로부터 수소를 제거하여 합성
④탄수 수가 16~18개 이상인 포화지방산이 원료
⑤동물에서 C9~C10(Δ9)의 불포화결합 형성 가능
but C12~C13(Δ12)와 C15~C16(Δ15)의 위치에서 불포화결합을 형성할 수 없음→리놀레산과 α-리놀레산은 합성 못함→이 두 지방산은 동
물에서 필수지방산임
⑥식물은 모든 불포화지방산을 생성할 수 있음
인지질 합성
①동물 : 활면소포체 표면
②식물 : 엽록체와 소포체 표면
③성장하고 있는 생물체에서는 세포막성분이 필요하므로 주로 인지질이 많이 합성되고 성장이 완료된 후는 주로 중서지방으로 합성됨
스테로이드 합성
①아세테이트 메발로네이트 경로
동물 : 간의 소포체, 콜레스테롤이 합성됨
식물 : 소포체, 스티그마스테롤
진균류(곰팡이) : 에르고스테롤 합성
②메틸 에리스리톨 인산염 경로 : 식물체에만 존재, 지베렐린, ABA, 카로티노이드 등 각종 생리활성 이소프레노이드 생성
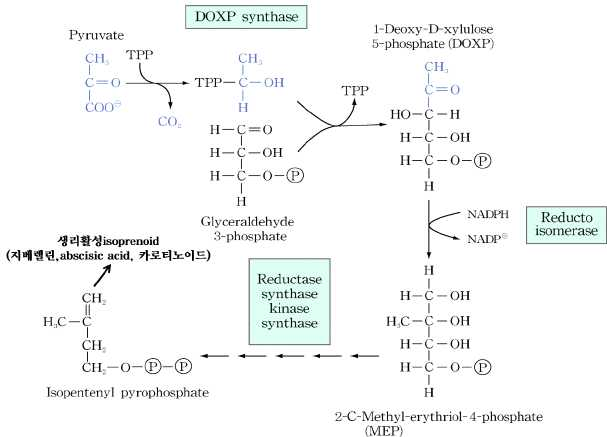
스테로이드-메틸 에리스리톨 인산염(MEP) 경로
(정리2)
⦁지방산은 동물에서는 에너지원으로 주로 이용되지만 식물에서는 발아하는 종자를 제외하고는 주로 물질 합성의 원료로 사용된다.
⦁ATP를 소모함으로써 열역학적인 불리함을 보전하고, ATP 분해 결과 생성된 이중인산이 신속하게 인산으로 전환됨으로써 지방산
CoA가 형성되는 정반응만이 일어날 수 있는 체계가 존재한다.
⦁프로피오닐 CoA가 이산화탄소의 낱소 1개를 공급받아 숙시닐 CoA로 전환된 후에 크랩스회로에서 에너지원으로 이용된다.
⦁불포화지방산은 그 지방산의 불포화결합이 탄소 1개를 공급받아 숙시닐 CoA로 전환된 후에 크렙스회로에서 에너지원으로 이용된다.
⦁식물은 필요한 모든 종류의 불포화지방산을 합성하지만 동물은 C₁₂~C₁₃ 사이, C₁₅~C₁₆ 사이의 불포화결합을 형성할 수 없기 때문이다.
⦁종자가 발아할 때에는 종자에 저장되어 있는 지질이 분해되어 에너지원과 물질 합성원료로 사용된다. 그러나 종자 외의 조직에서
는 광합성 및 탄수화물 에너지생산 대사에 의해 에너지가 공급되며, 지방질은 물질 합성의 원료로만 사용된다.
⦁지방산이 β-산화과정을 거치면 acetyl CoA, NADH, FADH₂가 각기 한 분자씩 생산된다. NADH, FADH₂는 전자전달계에서 ATP를
생산하고, acetyl CoA는 크렙스회로에서 이용된다.
⦁지방을 에너지원으로 과다하게 사용하면 acetyl CoA가 대량으로 생산된다. acetyl CoA가 크렙스회로에서 이용되기 위해서는 옥살
로아세트산이 필요한데, 옥살로아세트산이 부족하여 과다하게 생산되는 acteyl CoA의 일부만 크렙스회로에서 이용될 수 있다. 크
렙스회로에서 이용되지 못한 acetyl CoA는 케톤체를 생성한다.
⦁진핵세포에서는 C₁₆~C₁₈ 이상의 포화지방산에서만 불포화결합이 형성된다. 팔미트산은 C₁₆ 포화지방산이다.
⦁스테로이드 물질 합성경로는 acetate mevalonate pathway와 methyl eryhriol phosphate pathway인데 동물에서는 acetate
mevalonat pathway만이 존재하고, 식물에서는 acetate mevalonate pathway와 methyl erythriol phosphate pathway 모두
존재한다.
[인강연습문제]
1. 동물과 식물이 지방질을 이용하는 체계의 차이를 제시하시오.
☞식물은 필요한 만큼의 물질만을 합성하기 때문에 에너지원이 과다하여 지방으로 저장되는 일이 없다. 다만 종자의 경우에는 지방을
에너지원과 물질합성원료로 이용한다. 동물은 에너지원의 공급과 수요가 정확하게 일치하지 않기 때문에 과다하게 섭취된 에너지는 지
방으로 저장되고, 에너지원의 섭취가 부족하면 저장 되었던 지방이 에너지로 이용된다.
2. 지방산이 β-산화에 의해 분해 될 때 생산되는 에너지 관련 물질을 제시하시오.
☞ NADH, FADH2, acetyl CoA
3. 짝수 탄소의 지방산과 홀수탄소의 지방산이 β-산화에서 생산되는 물질의 차이를 제시하시오. ☞ 짝수 탄소 지방산 - NADH,
FADH2, acetyl CoA
☞ 홀수 탄소 지방산 - NADH, FADH2, acetyl CoA, succinyl CoA
4. 지방을 에너지원으로 과다하게 이용할 때 케톤증이 일어나는 이유는 무엇인가?
☞지방산이 과다하게 에너지원으로 사용되어 β-산화 결과 생산된 아세틸CoA가 에너지원으로 사용하지 못하기 때문이다.
5. 포유류에서 리놀레산과 리놀렌산이 필수지방산인 이유는 무엇인가?
☞포유류는 지방산의 C12-C13과 C15-C16의 불포화결합을 형성하지 못하기 때문이다.
[연습문제]
1. 동물과 식물에서 지방산의 이용 양태의 차이는 무엇인가?
→ 지방산은 동물에서는 에너지원으로 주로 이용되지만 식물에서는 발아하는 종자를 제외하고는 주로 물질 합성의 원료로 사용된다.
2. 지방산이 분해되기 위해서 미토콘드리아 안으로 이동할 때 지방산 CoA로 전환하는 반응은 에너지 흡수반응임에도 불구하고 지방산
CoA가 용이하게 형성될 수 있는 이유는 무엇인가?
→ ATP를 소모함으로써 열역학적인 불리함을 보전하고, ATP 분해 결과 생성된 이중 인산이 신속하게 인산으로 전환됨으로써 지방산
CoA가 형성되는 정반응만이 일어날 수 있는 체계가 존재한다.
3. 홀수탄소 지방산의 β-산화 결과 생성된 프로피오닐 CoA가 에너지원으로 이용되는 경로는 무엇인가?
→ 프로피오닐 CoA가 이산화탄소의 탄소 1개를 공급받아 숙시닐 CoA로 전환된 후에 크렙스회로에서 에너지원으로 이용된다.
4. 불포화지방산의 산화과정이 포화지방산의 산화과정과 다른 점은 무엇인가?
→ 불포화지방산은 그 지방산의 불포화결합이 β-산화과정의 중간물질인 에노일 CoA의 불포화결합의 형태 및 위치와 동일하게 전환되는
반응이 추가로 필요하다.
5. 동물에는 식물의 경우와는 달리 필수지방산의 개념이 존재하는 이유는 무엇인가?
→ 식물은 필요한 모든 종류의 불포화지방산을 합성하지만 동물은 C12∼C13 사이, C15∼C16 사이의 불포화결합을 형성할 수 없기 때문이다.

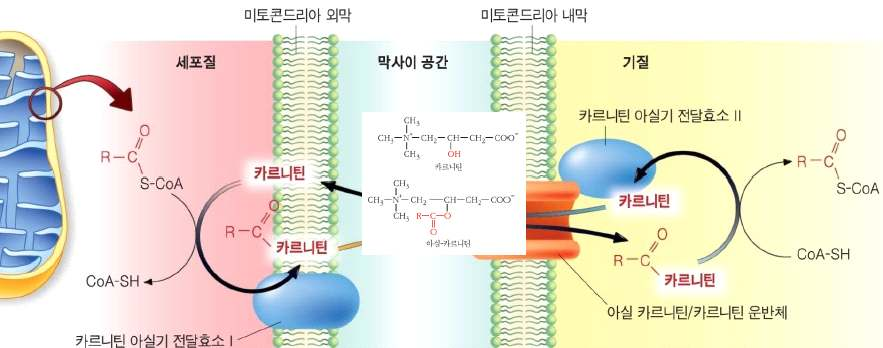
세포질: F atty acyl-CoA 합성 • 미토콘드리아 내: carnitine acyltransferase에 의해 기질로 전달
지방산 합성 과정

세포질: F atty acyl-CoA 합성 • 미토콘드리아 내: carnitine acyltransferase에 의해 기질로 전달
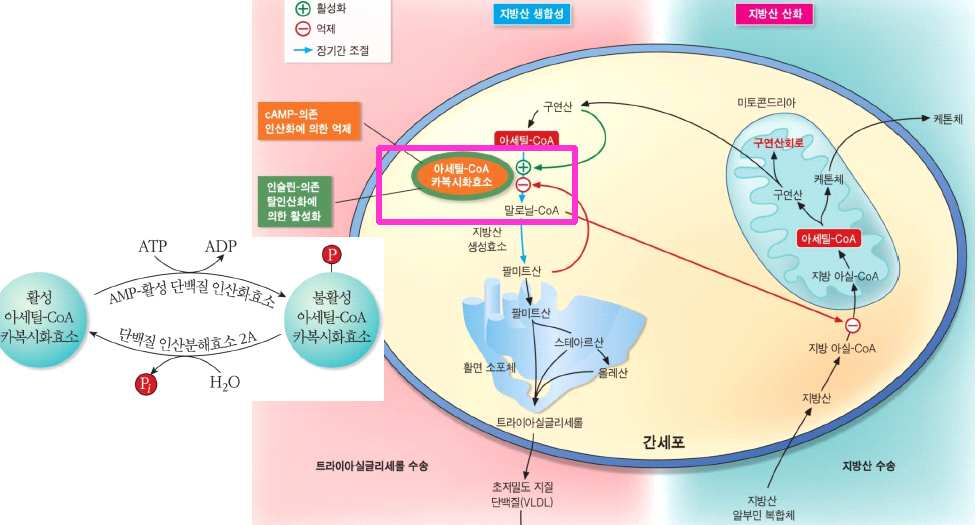
지방산 합성 조절아세 틸 CoA carboxylase: 말로 닐 CoA 생성말로 닐 CoA: 1) 팔 미 트 산 생성 = è T G 합성2) 카 르니틴 아 실 기 transferase I 억 제 = è 미토콘드리아로 들 어가는 작용 억 제 == è 산화 억 제
※ 부족하지만 글의 내용이 도움이 조금이라도 되셨다면, 단 1초만 부탁드려도 될까요? 로그인이 필요없는 하트♥(공감) 눌러서 블로그 운영에 힘을 부탁드립니다. 그럼 오늘도 행복한 하루 되십시오^^
'농업생물화학' 카테고리의 다른 글
| 농업직, 농학과, 화학, 방통대 등 농업생물화학 핵심 요점 정리 13. 핵산 대사 (19) | 2022.12.02 |
|---|---|
| 농업직, 농학과, 화학, 방통대 등 농업생물화학 핵심 요점 정리 12. 단백질 대사 (3) | 2022.12.02 |
| 농업직, 농학과, 화학, 방통대 등 농업생물화학 핵심 요점 정리 10. 탄수화물 대사 (5) | 2022.11.30 |
| 농업직, 농학과, 화학, 방통대 등 농업생물화학 핵심 요점 정리 9. 호르몬 (18) | 2022.11.27 |
| 농업직, 농학과, 화학, 방통대 등 농업생물화학 핵심 요점 정리 8. 효소반응 (9) | 2022.11.26 |