제 13장 핵산 대사
| ⦁개시코돈(initiation codon) : 단백질 합성의 개시를 지시하는 mRNA상의 3개의 ribonucleotide 조합이다. AUG가 개시코돈이다. 대장균의 경우 GUG가 개시코돈이 되는 경우가 있으나 매우 드물다. ⦁게놈 도서관(genome library) : 염색체 DNA를 불완전 가수분해한 후에 이를 유전자 운반체에 모두 결합시킨 것을 의미한다. 이 재조합 운반체들에 염색체 DNA에 존재하던 모든 유전자가 존재하기 때문에 게놈 도서관이라 한다. ⦁구조유전자(structual gene) : 유전자가 발현될 때 단백질의 아미노산 혹은 RNA로 나타나는 DNA의 부위 ⦁넌센스 코돈(nonsense codon) : 종결코돈은 해당하는 아미노산이 없기 때문에 넌센스 코돈이라고도 한다. ⦁발현 운반체(expression vector) : 유전자를 운반하여 숙주세포에서 발현케 하는 운반체 ⦁번역(translation) : mRNA 구조에 근거하여 polypeptide가 합성되는 과정 ⦁복제(replication) : DNA를 주형으로 하여 동일한 DNA가 합성되는 과정 ⦁복제기점(replication origin) : DNA가 복제될 때에 복제가 시작되는 부위 ⦁복제분기점(replication fork) : DNA가 복제될 때에는 DNA의 두 가닥 polydeoxyribonucleotide의 이중구조가 풀리면서 각 가닥이 복제되는데, 폴리는 지점을 복제분기점이라 한다. ⦁복제자(replicator) : 진핵생물의 복제기점이며 자율복제배열과 동의어이다. ⦁상보적 DNA(complememtary DNA: cDNA) : mRNA로부터 역전사에 의해 합성된 DNA ⦁선도가닥(leading strand) : DNA 복제에서 3′-5′의 polydeoxyribonucleotide를 주형으로 하여 합성되는 polydeoxyribonucleotide ⦁안티코돈(anticodon) : mRNA상의 코돈에 대응하는 상보적 염기로 구성된 ribonucleotide배열을 의미한다. tRNA에 존재한다. ⦁억제자(repessor) : 전사인자의 한 종류로서 RNA 중합효소에 의한 mRNA 합성을 억제한다. ⦁엑손(exon) : 진핵생물 구조유전자의 단백질 혹은 RNA에 대응하는 부위 ⦁오카자키 단편(Okazaki fragment) : 지연가닥이 복제되는 과정의 중간에 합성되는 DNA 단편 ⦁오퍼레이터(operator) : 억제자가 결합하는 DNA상의 조절유전자 부위 ⦁오페론(operon) : 생물학적 기능을 가지는 여러 개의 단백질 유전자의 발현을 함께 조절하는 체계가 존재하는 DNA의 부위 ⦁유전암호(genetic code) : mRNA상에서 1개의 아미노산에 해당하는 3개의 ribonucleotide 배열들에 관한 정보. 아미노산에 대한 각 암호를 코돈(codon)이라 한다. ⦁인트론(intron) : 진핵생물 구조유전자의 단백질 혹은 RNA에 대응하지 않아 번역되지 않는 부위 ⦁자율복제배열(autonomously replicating strand: ARS) : 진핵생물의 복제기점을 일컫는다. ⦁전사(transcription) : 발현될 유전자 구조에 근거하여 mRNA가 합성되는 과정 ⦁전사인자(transcription factor) : 비필수유전자의 발현을 조절하는 단백질 ⦁조절유전자(regulatory gene) : 유전자가 발현되는 것을 조절할 뿐이지 발현되어 생산되는 최종생산물의 유전정보에는 해당하지 않는 유전자 ⦁종결코돈(termination codon) : 단백질 합성의 종결을 지시하는 mRNA상의 3개의 ribonucleotide 조합이다. UAA, UAG, UGA가 종결코돈이다. ⦁증강자(enhancer) : 활성자가 결합하는 DNA상의 조절유전자 부위 ⦁지연가닥(lagging strand) : DNA 복제에서 5′-3′의 polydeoxyribonucleotide를 주형으로 하여 합성되는 DNA 단편 ⦁촉진유전자(promoter) : 전사의 개시를 조절하는 조절유전자의 일종 ⦁코스미드(cosmid) : plasmid에 bacteriophage λ의 packaging 및 숙주세포 감염에 필요한 유전자가 첨가된 것으로서 유전공학에서 분자량이 큰 유전자를 운반 할 수 있는 운반체이다. packaging 유전자라 함은 코스미드를 둘러싸서 bacteriophage λ의 형태로 되어 복제 혹은 발현될 숙주세포에 감염될 수 있도록 하는 유전자이다. ⦁클로닝 운반체(cloning vector) : 유전자를 운반하여 숙주세포에서 발현케 하는 운반체 ⦁클로닝(cloning) : 동일한 생물물질을 대량으로 복제, 생산하는 것을 의미한다. 일반적으로 동일한 DNA를 대량으로 생산하는 과정을 클로닝이라 한다. ⦁클론(clone) : 유전적으로 동일한 세포 혹은 생물물질을 의미한다. ⦁특이성 인자(specific factor) : 전사인자의 한 종류로서 RNA 중합효소가 promoter를 인식하는 특성을 변화하여 다른 promoter에 결합케 함으로써 발현되는 유전자의 종류를 변화시킨다. ⦁폴리솜(polysome) : mRNA상에 여러개의 리보솜이 결합되어 있는 상태를 말한다. 전사된 mRNA의 단백질들이 동시에 신속하게 합성되기 위한 현상이다. ⦁프리모솜(primosome) : DNA 복제에서 지연가닥의 합성에 관련되는 합성개시, 신장, 종결 등 여러 가지 기능을 가지는 효소의 복합체 ⦁프리브노(Pribnow) 상자 : 세균의 promoter 구조 중 공통된 배열 6개의 염기쌍을 칭하는 것으로서 (5′)TATAAT(3′)의 공통배열을 가지는데 유전자마다 다소 다를 수 있다. 이는 일반적으로 발현될 유전자 앞쪽 10 염기쌍 부위에 존재한다. ⦁플라스미드(plasmid) : 세포에 존해하는 염색체 외의 유전정보기구로서 스스로 복제될 수 있는 기능을 가지고 있다. 원핵세포와 효모에존재한다. plasmid에 존재하는 유전정보는 생명현상에 비필수적인 정보들이다. 세균의 plasmid는 유전공학에서 유전자 운반체로서 많이사용되며 겹환상의 구조를 가지고 있다. ⦁하우스키핑유전자(housekeeping gene) : 생명현상에 필수적인 정보를 가지고 있는 유전자로서 유전자 발현 조절없이 항상 일정량이 발현되는 유전자 ⦁호르몬 반응요소(hormone response element: HRE) : 유전자 발현을 억제 혹은 촉진하는 스테로이드계 호르몬이 결합하는 DNA상의 부위 ⦁활성자(activator) : 전사인자의 한 종류로서 활성자는 RNA 중합효소에 의한 mRNA 합성을 촉진한다. |
□ 핵산대사
ㅇ 유전자 보존(DNA 복제)
- 세포의 증식 : 세포수 증가, 세포 재생
- 번식 : 번식세포 형성
ㅇ 유전자 발현
- 전사 : mRNA, rRNA, tRNA 합성
- 번역 : 단백질 합성
- 발현 조절 : 필수 유전자, 비필수 유전자
ㅇ 유전자 재조합
- 유전자 분리, 형질 전환 ⦁유전자(gene) : 1개의 단백질 혹은 RNA합성에 필요한 DNA상의 정보.
→DNA상의 1개의 단백질 혹은 RNA정보에 해당하는 deoxyribonucleotide들의 결합순서.
⦁deoxyribonucleotide : DNA의 단량체(단위체). 보통 DNA를 구성하는 뉴클레오타이드가 단독으로 존재하는 형태. 각각의 디옥시리
보뉴클레오타이드는 질소염기, 디옥시리보(5탄당), 인산의 세부분으로 구성되어있다.
13.1. 유전자 구조 : 기능에 따라 구조유전자,조절유전자로 분류
1) 구조유전자
⦁원핵생물의 구조유전자는 모두 단백질 혹은 RNA에 해당하는 정보이나, 진핵생물체 구조유전자의 경우는 단백질(아미노산) 혹은 RNA에 대응하는 엑손(exon)과 대응하지 않아 번역되지 않는 인트론(intron) 부위로 구성되어 있다. 인트론을 개재배열(intervening sequence)이라고도 한다.
2) 조절유전자
⦁구조유전자가 단백질 혹은 RNA로 발현되는 것을 조절하는 유전자이다.
⦁단백질 합성을 위한 mRNA 전자를 지시하는 촉진유전(promoter)가 대표적인 조절유전자이며 구조유전자의 전산 개시점 앞쪽에
존재한다.
13.2. 유전암호

㉮ DNA의 유전정보에 근거해서 합성된 mRNA상의 1개의 아미노산에 대한 3
개의 ribo-nucleotide를 codon이라 하고, 코돈들 전체를 유전암호라 한다.
㉯ 유전암호는 mRNA의 5′→ 3′방향의 순서로 표기한다.
㉰ 유전암호는 아미노산 혹은 RNA에 대응하는 것 외에 mRNA의 합성(전사)
의 개시를 의미하는 개시코돈(AUG, GUG)과 전사의 종결을 의미하는 종결코
돈(UAA, UGA, UAG)이 있다. GUG가 개시코돈인 경우는 아주 드물다.
13.3. DNA 복제

⦁주형이 되는 어버이DNA의 복제기점의 이중나선이 풀리면서 각 주형가닥의 DNA에 상보적인 염
기를 가지는 데옥시리보뉴클레오티드가 결합되면서 DNA가 복제되어 어버이 DNA와 동일한 구조
의 DNA와 동일한 DNA가 2배로 증가하게 된다.
- 새로운 dNTPs의 PPi가 분해 제거되면서 5-인산이 신장해가고있는 DNA의 3말단 dNTPs의
3‘-OH에 결합하여 monophospho
-diester결합이 형성되어 새로운
DNA가 나간다. 즉 DNA의 복제는
5-3방향으로 진행된다.
※ DNA 복제과정에서 효소들의 작용
helicase→단일가닥 결합단백질→DNA
회전효소→primase→DNA 중합효소Ⅲ
→DNA 중합효소Ⅰ→DNA 연결효소
(ligase)

13.1.1 DNA 복제과정

(1) 개시
㉮ 복제기점에서 두 가닥의
polydeoxyribonucleotide가 풀
려 한 가닥씩으로 분리된다.
㉯ 풀린 각 가닥의 DNA에 DNA
결합단백질(single-strand DNA binding protein:SSB)이 결합하여 단일가
닥 DNA들이 다시 결합하지 못하게 한다.
㉰ DNA가 단일가닥으로 풀리면 회전하기 때문에 DNA가 뒤틀리는데 이러한
위상 스트레스(topological stress)는 DNA 선회효소(DNA gyrase; DNA
topoisomerase)가 해제한다.
(2) 신장
DNA 가닥이 풀리면 deoxyribonucleotide들이 결합되어 있는 방향이 한 가닥은 3′→ 5′의 것이고, 다른 가닥은 5′→ 3′이다.
DNA는 5′→ 3′방향으로 복제되어 나가기 때문에 3′→5′ 가닥을 주형으로 하면서 합성되는 가닥은 풀리는 방향으로 계속해서 DNA가 합성될 수 있다. 이렇게 합성되는 새로운 가닥을 선도가닥(leading strand)이라 한다. 그러나 5′→ 3′ 가닥을 주형으로 하는 경우에는 DNA가 풀리는 방향으로 DNA가 합성되지 못하고 반대방향으로 합성되어야 하기 때문에 DNA가 풀리면서 연속적으로 합성되지 못하고 이미 풀린 부위에 부분적으로 5′→ 3′방향으로 DNA 단편이 합성된 후에 단편들이 결합하여 새로운 DNA 가닥이 완성된다. 이러한 방법으로 완성되는 DNA 가닥을 지연가닥(lagging strand)이라 한다.
① 선도가닥

㉮ 3′→ 5′의 DNA를 주형으로 하여 상보적인 염기를 가지는 deoxyribonucleotide가 결합하면서 새로운 DNA 가닥이 5′→ 3′방향으로 연속적으로 복제된다.
㉯ 주형 DNA에 상보적인 염기를 가지는 deoxyribonucleotide가 결합을 시작하기 위해서는 기초물질이 필요하다. 이 기초물질을 시발물질(RNA primer)이라 한다. 시발물질은 시발효소(primase)가 제조한 것으로서 10∼60개의 ribonucleotide로 구성되어 있다. 시발물질의 3′말단에 상보적인 염기를 가지는 deoxyribonucleotide의 5′인산이 DNA 중합효소Ⅲ에 의해 결합하면서 선도가닥이 합성된다. 합성이 완료된 후에 시발물질이 분해, 제거되고 그 자리에 주형
가닥의 상보적 염기를 가지는 deoxyribonucleotide가 결합한다.
② 지연가닥

㉮ 지연가닥은 복제분기점이 이동하는 방향과 반대방향(3′→5′)으로 새로운 DNA가 합성된다.
㉯ DNA가 풀리면 개시, 신장, 종결에 관련된 모든 효소의 기능을 가지는 primosome이 풀린 5′→ 3′을 주
형으로 하여 5′→ 3′방향으로 수시로 이동하면서 RNA primer를 합성하여 결합시킨다. DNA 중합효
소Ⅲ가 RNA primer의 3′말단에 주형DNA와 상보적인 염기를 가지는 deoxyribonucleotide를 결합하여
새로운 DNA 단편을 여러 군데 형성한다. 이 불연속적인 DNA 단편을 오카자키 단편(Okazaki fragment, DNA의 지연가닥 복제시 형성되는 단편)
이라 한다. 오카자키 단편들 간의 연결되지 않은 부위를 새김눈(nick)이라 하고 새김눈은 DNA 연결효
소에 의해 연결되어 새로운 DNA 가닥이 완성된다.
(3) 종결
복제가 완료된 DNA는 주형 DNA와 엉켜 있다. 위상이성질화효소(topoisomerase) IV가 한쪽 염색체의 두 DNA 가닥을 일시적으로 분해하여 엉킨 DNA가 빠져나와 분리되도록 한다.
13.3.2 DNA복제 오류 수정
복제의 오류를 수정하는 체계가 있기 때문에 유전정보가 매우 정확하게 복제되어 보존될 수 있다.(ex:돌연변이)
(1) 잘못된 상보적 염기결합 수복
복제되는 과정에서 잘못 결합된 deoxyribonucleotide를 복제 2∼3초 후에 제거하고 옳은 deoxyribonucleotide로 수복한다.
(2) 염기제거 수복
복제된 DNA의 염기가 탈아미노화, 알킬화, 인접한 피리미딘염기 간의 결합등에 의해 손상이 있으면 염기만을 제거하여 수복한다.
(3) 손상 부위 디옥시리보뉴클레오티드 절제, 수복
DNA 구조에 큰 변화가 있을 정도로 변화, 손상된 부위가 있는 경우 해당 부위 뿐만 아니라 부근의 deoxyribonucleotide들도 함께 제거한 후 수복한다.
(4) 직접 수복
손상된 염기가 있는 경우 염기를 제거하지 않은 상태에서 손상된 부위만을 수복하는 경우가 있다.
※ DNA유전암호의 전사(transcription)와 번역(해독,translation)

⑴ DNA의 유전정보를 전달하는 RNA를 mRNA라고 하며 DNA의 유전정보에 따라 mRNA를 합성하는 과정을 전사하고 한다. 진핵생물의 경우 전사는 핵 속에서 일어난다
⑵ 전사된 mRNA는 핵공을 통해 세포질로 빠져나와 리보솜과 결합한 후 단
백질 합성에 이용된다. mRNA에 저장되어 있는 정보를 이용하여 단백질
을 합성하는 과정을 번역이라고한다. 번역은 세포질에서 일어난다.
① DNA 유전암호(코드) ② mRNA의 전사(코돈) ③ tRNA의 운반(안티코돈)
④ 리보솜에서 단백질 합성(번역)
⑶(생명 중심설) DNA에 저장되어 있는 유전정보는 직접 단백질
합성에 관여하지 않고 DNA정보가 RNA로 전사된 후에 RNA에 의해서 단백질 합성이 일어난다.

13.4. 전사 : DNA유전정보 ⇒ RNA합성
ㅇ 전사 생산물 (mRNA : 단백질 합성, rRNA : ribosome 구성, tRNA : 아미노산 운반)
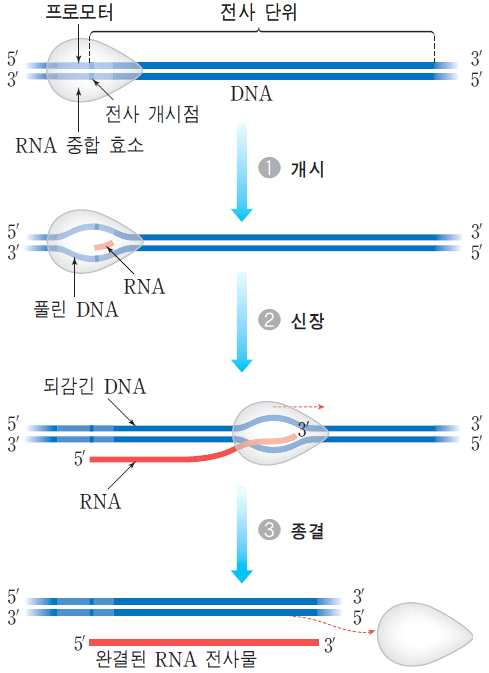

전사란 DNA에 존재하는 유전정보가 발현되기 위해 DNA의 정
보에 근거하여 RNA가 합성되는 과정이며, 합성되는 RNA는
mRNA, rRNA, tRNA이다. mRNA는 합성되는 단백질의 아미
노산 구성 및 순서와 관련한 정보가 전사된 RNA이고 rRNA와
tRNA는 mRNA에 전사된 유전정보에 따라 단백질이 합성되는
것을 지원하는 기능을 가진다.
1) 전사 개시

→DNA의 프로모터(RNA 중합효소 부착하는 장소)에 RNA 중합
효소와 전사인자가 결합하여 전사개시 복합체를 형성하면서 염
기사이의 수소결합을 절단함으로ㅆ DNA의 이중가닥이 풀어진
다.
㉮ DNA상에서 발현되어야 할 부위의 정보에 근거하여 mRNA
가 합성되는 것을 전사라 하는데 전사의 개시를 조절하는 유전
자를 촉진유전자(promoter)라 한다. RNA를 합성하는 RNA 중
합효소가 촉진유전자에 결합할 수 있어야 전사가 시작된다.
RNA 중합효소가 촉진유전자에 결합할 수 있는 환경은 전사인
자, 유도물질 등과 같은 유전자 조절체계에 의해 이루어진다.
㉯ 대장균 촉진인자는 5′→ 3′ DNA 가닥의 발현될 유전자 5′전
방 -10bp 부위와 -35bp에 공통배열이 존재한다. -10bp 부위
에 있는 5′TATAAT3′의 공통배열은 이를 발견한 David
Pribnow의 이름을 따서 프리브노(Pribnow) 상자라고 한다.
-35bp 부위의 공통배열은 5′TTGACA3′이다.
㉰ 진핵생물 촉진인자는 -30bp 부위, +1bp 부위의 공통적인
배열을 가지는 곳과 공통성이 없는 조절 부위가 존재한다.
-30bp 부위와 +1bp 부위의 공통구조는 각기 5′TATAAAA
3′, 5′YYAN YY3′이며 각각 TATA 상자, Inr 서열(Initiator,
개시자, 촉진유전자의 일부)이라 한다. Inr 서열의 N의 위치에
는 모든 디옥시리보뉴클레오티드가 가능하고 Y위치에는 피리
미딘(시토신, 티민) 디옥시리보뉴클레오티드가 가능하다.
2) 전사 (RNA 합성)

㉮ RNA 중합효소가 전사될 DNA에 결합한 후 촉진유전자로 이
동하여 결합하면 DNA상의 염기쌍이 풀리면서 전사될 부위의
주형사슬(-)의 deoxyribonucleotide가 노출되면서 mRNA가
합성되기 시작한다.
주형사슬은 3′→ 5′의 polydeoxyribonucleotide이며, 5′→ 3′
polydeoxyribonucleotide는 비주형사슬(+) 혹은 코딩사슬이라 한다.
㉯ DNA에 상보적인 염기를 가진 ribonucleotide가 결합하고 RNA 중합효소가 ribonucleotide들을 결합한다. RNA 합성에는 DNA가 복제될 때와는 달리 시발물질(primer)이 필요하지 않다.
3) 전사 종결
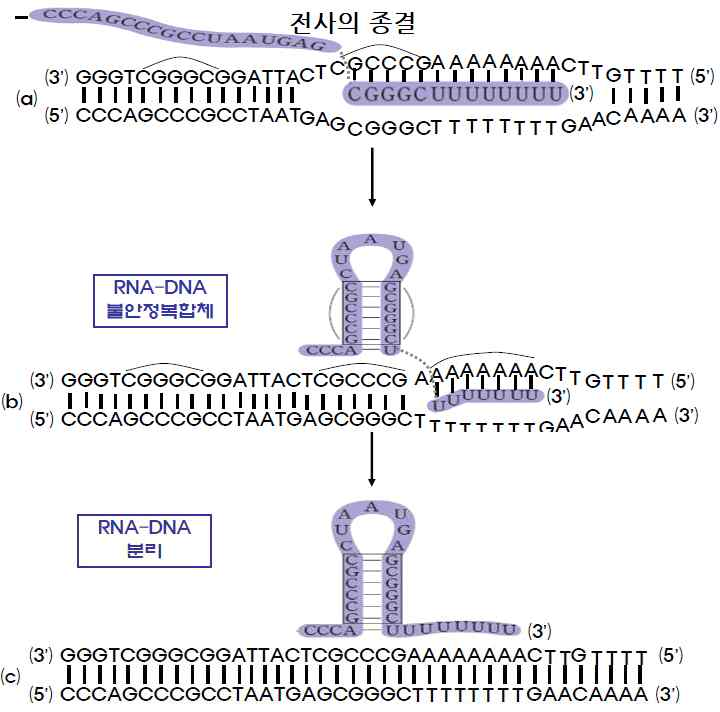
㉮ 전사는 주형 DNA상에 존재하는 RNA 중합효소가 DNA로부터 분리되어야 할
구조가 나타날 때까지 계속된 후에 RNA 중합효소가 DNA와 분리하면서 전사
가 종결된다.
㉯ 전사가 종결될 주형 DNA상의 말단 부위에 자기 상보적 배열이 존재하고 그
이후에 아데닌염기(A)가 집단으로 존재하면 합성된 RNA는 머리핀구조를 가
지면서 후미에 우라실염기(U)의 deoxyribonucleotide를 집단적으로 가진다.
4) 전사오류
㉮ RNA 중합효소는 DNA 중합효소의 복제오류 수정 기능에 해당하는 3′→ 5′
TA exonuclease 활성이 없어 전사의 오류 확률은 1/104∼1/105로서 높은
편이다. 그러나 RNA는 생산 후에 기능을 마치면 분해되기 때문에 전사오류
는 유전정보 보존성에 영향을 미치지 않는다.
㉯ RNA를 게놈으로 가지고 있는 바이러스에서는 RNA 구조에 근거하여 DNA를
합성하는 역전사효소는 RNA 중합효소와 마찬가지로 3′→ 5′방향으로 오류를
수정하는 exonuclease 활성이 없으며 DNA 합성에서 1/2,000의 오류가 있
어 오류빈도가 매우 높다. 이는 변이 병원성 바이러스가 빈번하게 발생하는
원인이다.

5) RNA구조 완성 (DNA⇒RNA)
v mRNA
- 진핵세포 : splicing, 5’cap, poly(A) 꼬리구조
- 원핵세포 : 5’cap, poly(A) 꼬리구조
v rRNA
- 진핵세포 : 전사(45S), 메틸화, 절단(18S, 28S, 5.8S)
- 원핵세포 : 전사(30S, tRNA포함), 메틸화, 절단(16S, 23S, 5S)
v tRNA
- 5’ 및 3’ 말단 불필요부위 제거, splicing(진핵세포)
3’말단 5’CCA3’ 첨가, 메틸화, 탈아미노기 반응
진핵생물은 원핵생물에 비해 유전자의 수가 많고 유전체의 크기도 매우 크다. 하나의 수정란에서 만들어진 세포들도 발현되는 유전
자의 종류와 수준, 시기가 달라 서로 다른 세포로 분화된다. 전사와 번역의 장소가 핵과 세포질로 구분되어 있고, RNA의 가공 과
정, 단백질의 변형 과정을 거친다. 이러한 많은 단계에서 유전자 발현이 조절된다. 진핵 세포의 경우 전사는 핵에서, 번역은 세포질
에서 일어나기 때문에 전사 결과 처음 만들어진 RNA가 가공 과정을 거쳐 성숙한 mRNA로 된다. mRNA는 핵공을 통해 세포질로
수송되어 번역에 이용된 후 분해되며, 합성된 단백질은 변형 과정을 거친다. → 대부분의 단백질은 번역 후 변형 과정을 거쳐 특정
기능을 가진다. 진핵생물의 DNA가 단백질로 발현되기까지는 전사 조절, 전사 후 조절, 번역 조절, 번역 후 조절 등 여러 단계에서
조절될 수 있다.
-원핵 세포 : DNA가 세포질에 있어서 전사와 번역이 동시에 일어난다.
-진핵 세포 : DNA가 핵 속에 있어서 전사는 핵 속에서, 번역은 세포질에서 일어난다.
(1) mRNA
① 진핵세포 : 모든 DNA에는 단백질로 번역되지 않는 intron부위가 다수 존재하기 때문에 intron이 제거되고 엑손 부위의 정
보에 의해 구성된 구조로 완성되어야 한다.(발현되지 않는 intron이 존재한다)
㉮ 원핵세포의 1차 전사물의 정보는 모두 합성될 단백질의 아미노산과 일치하지만 진핵세포의 경우는 불필요한 인트론 부위가
제거되어야 한다. 이를 splicing이라 한다.
㉯ 전사된 mRNA의 5′말단 부위에 7-methylguanosine 잔기가 삼인산에 의해 결합되어 있는데 이를 5′cap이라 한다. 이는
단백질 합성(번역) 시 mRNA가 리보솜에 결합하는 것을 도와주고 mRNA가 분해하는 것을 방지한다.
㉰ mRNA가 합성된 후 3′말단에 여러 개의 아데닐산이 결합하여 poly A 꼬리가 형성된다. 이는 mRNA의 분해를 방지한다.
② 원핵세포(세균) : DNA구조는 합성될 RNA 혹은 단백질의 아미노산배열과 일반적으로 일치한다
mRNA가 합성된 후 3′말단에 여러 개의 아데닐산이 결합하여 poly A 꼬리가 형성되는데 이는 진핵세포와는 달리 mRNA의 분해를 촉진한다.
(2) rRNA
① 진핵세포 : 45S 크기로 전사된 후에 필요한 부위가 메틸화하여 18S, 28S, 5.8S 부위로 절단되면서 불필요한 부위는 제거된다.
② 원핵세포
30S rRNA로 전사된 후에 필요한 부위만이 메틸화하여 tRNA, rRNA의 16S, 23S, 5S의 단위체로 분리되고 불필요한 부위는 제
거된다. 30S rRNA에 tRNA가 함께 포함되어 있으며 전사 후 가공과정에서 분리된다.
(3) tRNA
㉮ tRNA는 5′말단과 3′말단에 불필요한 부위가 포함되어 전사되기 때문에 이
들이 제거된다. 진핵세포의 경우에는 인트론이 존재하기도 하며 이는 가공과정
에서 제거된다.
㉯ 3‘말단에 5′CCA3′가 결합하며, 새로이 형성된 3′말단의 아데닐산(A)의
-OH기에 운반될 아미노산이 결합한다.
㉰ tRNA의 염기 중 일부는 메틸화, 탈아미노반응, 환원 등에 의해 변형된다.

3.5. 번역(합성) DNA⇒RNA⇒단백질
mRNA에 전사되어 존재하는 유전정보에 근거하여 단백질이 합성되는 것을 번
역(translation)이라 한다. 단백질 합성은 유전정보가 발현되는 과정이며, 세포
내 생합성에 소모되는 에너지 중 90%가 단백질 합성에 이용된다. 번역은 아미
노산 활성화, 개시(initiation), 신장(elongation), 종결(termination), 단백질구
조 완성의 단계로 일어난다.
1) 아미노산 활성화
단백질이 합성되기 위해서는 tRNA가 아미노산을 ribosome에 결합되어 있는
mRNA로 운반하여야 한다. 이때 아미노산이 tRNA에 결합하기 위해서는 ATP
와 반응하여 5′-aminoacyl adenylate(aminoacyl-AMP)로 활성화해야 한다.
2) 개시
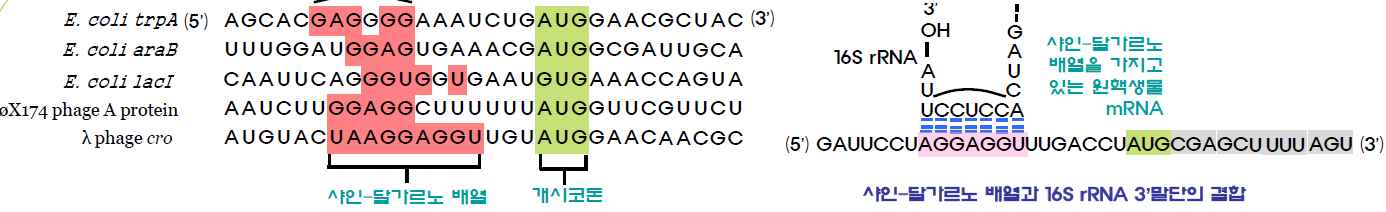
㉮ mRNA와 ribosome이 결합한 후 mRNA상의 개시코돈인 AUG에 첫 번째 아미노산이 tRNA에 의해 운반되어 결합한다. AUG는
개시코돈인 동시에 methionine에 해당하는 유전암호이다.
㉯ 대장균(원핵)에서는 개시코돈인 AUG에는 mRNA 중간에 존재하는 AUG와는 달리 formyl화되어 있는 methionine과 tRNA 복합체인 fMet-tRNAfMet만이 결합할 수 있고 formyl화되어 있지 않은 methionine과 tRNA 복합체인 Met-tRNAMet은 결합하지 못하는데 이는 샤인-달가르노 배열이 개시코돈 전방 근처에 존재하기 때문이
다. 즉 5′AUG가 polypeptide 첫 번째 아미노산인 N-formyl -methionine에 해당하는 유전암호로 인식되는가 혹은 polypeptide 중간의 methionine에 해당하는 유전암호로 인식되는가는 샤인-달가
르노 배열이 mRNA상의 AUG 유전암호로부터 5′방향으로 8∼13 염기 부근에 존재하는가 여부에 의해 결정된다.
㉰ 진핵세포의 개시코돈 AUG 부근에 샤인-달가르노 배열과 유사한 부위가 존재하지 않기 때문에 리보솜이 mRNA의 5′말단으로부터 개시코돈을 찾아 가야 한다. 진핵세포에서 mRNA의 개시코돈(AUG)에는 메티
오닌이 운반된다. 그러나 진핵생물의 미토콘드리아와 엽록체에서 단백질이 합성될 때에는 개시코돈에 N-formylmethionine이 운반된다.

3) 신장
㉮ tRNA가 mRNA상의 유전암호에 따라 해당하는 아미노산을 운반하고, 아미노산 간에 peptide결합이 형성되면서 polypeptide가 합성된다.
㉯ mRNA상의 유전암호에 따라 폴리펩티드상에서 아미노산이 정확한순서로 결합하는 것은 아미노산을 운반하는 tRNA의 안티코돈이 mRNA의 코돈에 정확하게 결합하기 때문이다.
4) 종결
polypeptide의 신장은 mRNA의 종결코돈(UAA, UAG, UGA)에 도달하기까지 계속된다. mRNA의 종결코돈에 도달하면 방출인자 (releas factors: RF)가 polypeptide를 tRNA로부터 분리하고, 리보솜도 단위체로 분리하여 폴리펩티드 합성이 종결된다.
5) 단백질구조 완성
- 입체 구조 : 2차, 3차, 4차 구조 형성 → 생물학적 기능
- 5’말단 : 두번째 아미노산(ala, ser, thr)
- Oligosaccharide : o-OH-ser, thr N-liked- asp-X-ser(thr)
- 인산화 : ser, thr, tyr
(1) 입체구조
적절한 2차, 3차, 4차구조의 입체구조가 완성되어 단백질이 필요로 하는 생
리적 기능을 가지게 된다.
(2) 아미노기말단의 formylmethionine
원핵세포에서는 두 번째 아미노산이 alanine, serine, threonine과 같이 R-
부위가 짧은 아미노산이면 첫 번째 formylmethionine이 분해, 제거될 수
있다.
(3) 신호배열
아미노기말단의 15∼30 아미노산이 세포 내에서 최종 장소로 이동하는 신
호배열인 경우 최종 장소로 이동한 후에 분해, 제거된다.
(4) 인산화
serine, threonine, tyrosine에 인산이 결합될 수 있다. 우유단백질이
casein의 serine들이 인산화되어 있는데 인산에 Ca이 결합되어 있다.
(5) oligosaccharide의 결합
진핵생물의 경우 serine, threonine의 -OH기에 oligosaccharide가 산소
(O)를 매개로 하여 결합할 수 있다. asparagine- X-serine(혹은
threonine)의 순서로 아미노산이 배열되어 있을 때 asparagine의 N를 매
개로 하여 oligosaccharide가 결합한다.
※ 단백질 합성과정-번역

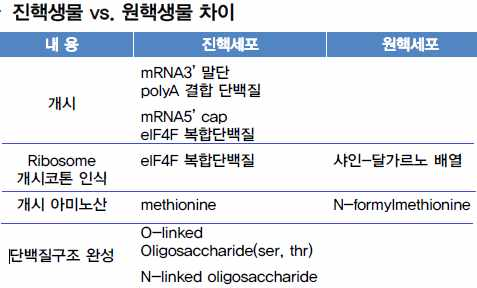
1. 단백질 합성 과정
세포질에서 mRNA의 코돈에 따라 아미노산을 붙여 특정한 단백질을 합성하는 과정
을 번역이라고 한다.
단백질 합성은 개시, 신장, 종결의 3단계로 이루어진다.
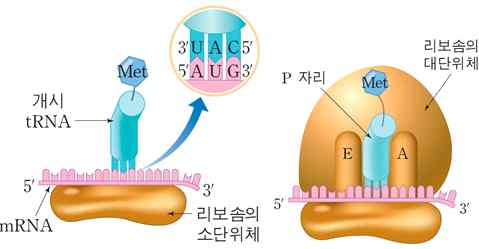
2. 번역의 개시
mRNA가 개시 tRNA 및 리보솜의 소단위체와 결합하고, 여기에 리보솜의 대단위체
가 부착하여 개시 복합체를 형성한다.
ⓐ 개시 tRNA의 결합 : 리보솜 소단위체가 mRNA와 결합하고, 개시 tRNA의 안티코돈
이 mRNA의 개시 코돈에 상보적으로 결합한다.
ⓑ 개시 복합체 : 리보솜 대단위체가 결합하여 완전한 리보솜을 만든다. 개시 tRNA는
리보솜의 P자리에 위치하며 A자리에는 다음 아미노산을 가진 tRNA가 들어온다.
3. 신장 : 폴리텝타이드 사슬의 신장(elongation)
tRNA가 운반한 아미노산이 폴리펩타이드 사슬의 말단에 하나씩 결합된다.
ⓒ 다음 tRNA 결합 : A자리의 mRNA 코돈과 상보적인 안티코돈을 가지고 있는 다음 tRNA가 A자리에 들어와 mRNA와 결합한다.(코돈의 인식)
ⓓ 폴리펩타이드 결합 : P자리의 tRNA에 붙어 있던 아미노산과 새로 운반되어 온 A자리의 아미노산 사이에 펩타이드 결합이 형성된다.
ⓔ 리보솜의 이동 : 리보솜이 mRNA의 3´ 말단 방향으로 이동하면 A자리에 있던 tRNA는 P자리로 이동하고, P자리에 있던 tRNA는 E자리로 이동
한 후 떨어져 나간다. (ⓒ ~ ⓔ 과정이 반복되면서 폴리펩타이드가 길어진다.)
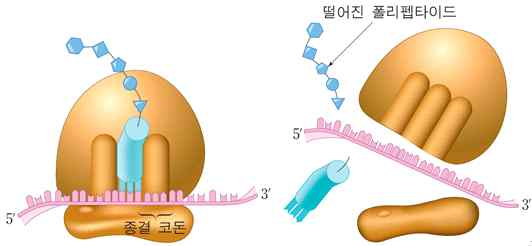
4. 종결
리보솜이 mRNA의 종결 코돈이 도달하면 tRNA 대신 방출 인자라고 불리는 단백질이 A자리에 결합한다. 방출 인자는 tRNA에 결합되어 있는 폴리펩타이드 사슬을 tRNA로부터 분리하여 방출시킨다.
ⓕ 단백질 합성 종결 : A자리에 mRNA의 종결 코돈이 도달하면 tRNA 대신 방출 인자라고 불리는 단백질이 A자리에 결합하며 폴리펩타이드의 신장이 중단된다.
ⓖ 리보솜의 분리 : 폴리펩타이드 사슬이 방출되고 리보솜의 두 단위체와 mRNA 등이 분리된다.

13.6. 유전정보 발현 조절
ㅇ 구성적 유전자
- RNA 중합효소와 촉진유전자 친화도
ㅇ 비필수 유전자
- 유도(induction)
- 전사인자(transcription factor) : 특이성 인자, 억제자(-), 촉진자(+)
1) 구성적 유전자 발현 조절(constitutive gene expression) : 조절없이 일정한 양이 항상 발현되는 현상
-세포에 필수적인 유전정보인 Housekeeping gene은 mRNA를 합성하는 RNA 중합효소와 DNA의 promoter 부위와의 친화성에 의하여 생산량이 조절된다. promoter가 다른 promoter들의 공통구조와 유사한 구조를 가질수록 promoter(촉진자)에 RNA 중합
효소가 잘 결합하여 mRNA가 다량으로 전사된다.
※Housekeeping gene:유전자에 존재하는 유전정보 중 세포의 생명현상에 필수적인 것은 일정량이 조절 없이 항상 발현되는 유전자.
2) 비필수유전자 발현 조절

(1) 전사인자
비필수유전자의 발현은 단백질인 전사인자(transcription factor)에 의해 조절되며, 기능에 따라 특이성 인자(specific factor), 억제자(repressor), 활성자(activator)로 분류한다. 전사인자가 결합하는 DNA상의 위치는 촉진유전자 내부, 전방, 후방에 존재하며, 촉진유전자 전방 수천 염기쌍의 거리의 부위에 결합하기도 한다.
세균에서는 전사인자를 필요로 하는 RNA 중합효소라 해도 전사인자가 없는 상태에서 어느 정도 촉진유전자에 결합할 수 있지만, 진핵세포에서는 전사의 중심 기능을 담당하는 RNA 중합효소Ⅱ는 전사인자가 없는 상태에서는 촉진유전자와 결합하지 않는다. 그 러므로 진핵세포에서는 전사인자가 유전자 발현에 반드시 필요하다.
① 특이성 인자
특별한 환경에서 RNA 중합효소가 promoter를 인식하는 특성을 변화시켜 다른
promoter에 결합케 함으로써 발현되는 유전자의 종류를 변화시킨다.
② 억제자
억제자는 RNA 중합효소에 의한 mRNA 합성을 억제하며, 이를 부(負)조절(negative
regulation)이라 한다. 억제자가 결합하는 DNA상의 부위를 오퍼레이터(operator)라 한
다.
③ 활성자(촉진자)
활성자는 RNA 중합효소에 의한 mRNA 합성을 촉진하며, 이를 양(陽)조절(positive
regulation)이라 한다. 활성자가 결합하는 DNA상의 부위를 증강자(enhancer)라 한다.
(2) 오페론
㉮ 세균에서는 특정 형질에 관련하는 여러 개의 유전자가 나란히 집단적으로 존재하면서 하나의 조절체계에 의해 각 유전자들의 발
현이 함께 조절되는 경우가 있다. 이와 같은 여러 개의 기능적 단백질을 합성하는 데 필요한 유전자의 발현을 동시에 조절하는 영
역을 오페론(operon)이라 한다. 특정 아미노산 합성에 필요한 여러 효소들의 유전자 발현은 일반적으로 하나의 오페론에 의해 조
절된다.
㉯ 세균과 하등 진핵생물의 세포에서는 이와 같은 오페론이 다수 발견되었다. 그러나 고등 진핵생물의 세포에서는 오페론에서와 같이 유전자가 서로 연계되어 전사되지 않고 개별적으로 전사된다.
유전자 발현 조절
하나의 개체에 들어 있는 세포들은 모두 같은 유전자를 가지고 있다. 그러나 세포에 따라 유전자의 발현 수준이나 시기가 달라서 세포는 다양한 형태와 크기, 기능을 가진다. 다양하고 많은 수의 유전자의 발현이 조절되지 않고 모든 유전자가 항상 발현된다면 정상적인 세포로 발현될 수 없을 것이다. 유전자 발현의 조절 과정은 원핵생물과 진핵생물에서 그 양상이 다르게 나타난다. 원핵생물에서 유전자 발현의 조절은 주로 전사 단계에서 일어난다. 서로 연
관된 기능을 수행하는 여러 개의 유전자가 하나의 전사 단위로 묶여 이들의 발현을 한 번에 촉진, 억제할 수 있도록 하는 조절 과정이 존재한다. 진핵생물의 유전자 발현 조절은 전사 조절, 전사 후 조절, 번역 조절, 번역 후 조절 등 다양한 단계에서 조절 과정을 거친다.
<원핵생물 유전자 발현조절>

○ 오페론
1. 오페론
원핵세포의 DNA에서 프로모터와 작동 유전
자 및 기능적으로 연관되어 있는 유전자들
이 모여 있는 유전자 집단.
오페론의 구성 : 프로모터 + 작동 유전자 +
구조 유전자(군)
오페론은 원핵생물의 유전자 발현을 조절하
는 유전자 집단으로 진핵생물에는 존재하
지 않는다.
2. 오페론 유전자
프로모터 : RNA 중합 효소가 결합하여 전사
가 시작되는 DNA 부위
※ 원핵생물의 RNA 중합 효소 : 주형
DNA 가닥의 염기에 상보적인 염기를 결합
해 단일 가닥의 RNA를 합성하는 효소
작동 유전자(작동 부위):전사를 조절하는
스위치 역할을 하며, 억제 물질이 활성화되어 결합하는 DNA 부위
구조 유전자(암호화 부위):단백질 합성에 대한 유전 정보를 저장하고 있는 DNA 부위로, mRNA로 전사되어 리보솜에 유전 정보를 전달한다.
○ 대장균의 젖당 오페론
대장균은 포도당이 있을 때에는 포도당을 섭취하여 증식하지만, 포도당이 없고 젖당만 있을 때에는 젖당을 섭취하여 증식한다.
▶ 대장균의 에너지원
대장균은 포도당과 젖당을 모두 에너지원으로 사용할 수 있다. 젖당은 포도당과 갈락토스가 결합한 이당류이기 때문에 대장균이 젖당을 에너지원으로 이용하기 위해서는 먼저 젖당을 포도당과 갈락토스로 분해해야 한다. 젖당을 이용하기 위해서는 세 가지 효소를 생산해야 하는데, 이 세 가지 효소의 유전자 발현 과정은 젖당의 유무에 따라 조절된다. 젖당 오페론 : 대장균에서 젖당 분해 효소 생산을 조절하는 오페론이다.
○ 젖당 오페론의 발현 조절 과정
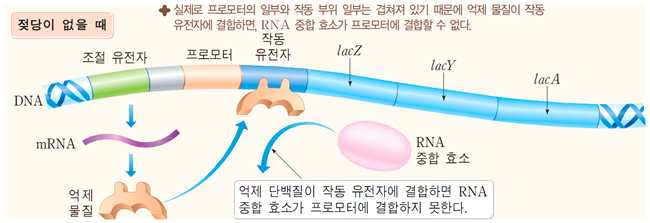

1. 젖당이 없을 때 : 구조 유전자의 전사가 억제된
다.
젖당 오페론이 아닌 조절 유전자에 의해 발현된 억제
물질이 프로모터의 작동 부위에 결합하여 RNA 중합
효소가 프로모터에 부착하는 것을 방해한다. 구조 유
전자의 전사가 일어나지 않아 젖당 분해에 필요한
효소를 합성하지 못한다.
2. 포도당이 없고 젖당만 있을 때 : 구조 유전자의
전사가 촉진된다.
조절 유전자에 의해 생산된 억제 물질이 젖당 유
도체와 결합하여 비활성화되므로 작동 부위에 결
합하지 못한다. RNA 중합 효소가 프로모터에 결
합하여 구조 유전자의 전사가 일어나게 된다.
3. 구조 유전자에 의해 발현된 단백질
β 갈락토시데이스 : 이당류인 젖당을 단당류인
포도당과 갈락토스로 분해한다.
투과 효소 : 젖당을 세포 안으로 운반시킨다.
아세틸 전이 효소 : 젖당을 분해할 때 보조 기
능을 한다.
4. 젖당 오페론의 활성화 시기
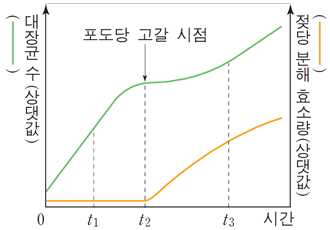
포도당과 젖당이 함께 들어 있는 배지에 대장균을
넣고, 시간에 따른 대장균의 수와 대장균 내의 젖당 분해 효소량을 나타낸 것이다.
t1시기의 대장균은 배지의 포도당을 에너지원으로 이용한다. 따라서 젖당 오페론이 작동하지 않는다. →
대장균은 포도당과 젖당이 함께 있을 때 포도당만 이용한다.
t2 시기에 포도당이 고갈되면 대장균에서 젖당을 에너지원으로 이용하기 위해 젖당 오페론이 활발하게
작동하기 시작한다. 이 시점부터 젖당 분해 효소량이 증가하기 시작한다.
t3시기의 대장균은 배지의 젖당을 흡수한 후 포도당으로 분해하여 에너지원으로 이용한다. 젖당 오페론
이 활발하게 작동한다.
<진핵생물의 유전자 발현>

진핵생물은 원핵생물에 비해 유전자의 수가 많고 유전체의 크기도 매우 크다. 하나의 수정란에서 만들
어진 세포들도 발현되는 유전자의 종류와 수준, 시기가 달라 서로 다른 세포로 분화된다. 전사와 번역
의 장소가 핵과 세포질로 구분되어 있고, RNA의 가공 과정, 단
백질의 변형 과정을 거친다. 이러한 많은 단계에서 유전자 발
현이 조절된다. 진핵 세포의 경우 전사는 핵에서, 번역은 세포
질에서 일어나기 때문에 전사 결과 처음 만들어진 RNA가 가공
과정을 거쳐 성숙한 mRNA로 된다. mRNA는 핵공을 통해 세포
질로 수송되어 번역에 이용된 후 분해되며, 합성된 단백질은
변형 과정을 거친다. → 대부분의 단백질은 번역 후 변형 과정
을 거쳐 특정 기능을 가진다. 진핵생물의 DNA가 단백질로 발
현되기까지는 전사 조절, 전사 후 조절, 번역 조절, 번역 후 조
절 등 여러 단계에서 조절될 수 있다.
◇ 원핵생물과 진핵생물
원핵 세포 : DNA가 세포질에 있어서 전사와 번역이 동시에
일어난다.
진핵 세포 : DNA가 핵 속에 있어서 전사는 핵 속에서, 번역은
세포질에서 일어난다.
○ 진핵생물의 유전자 발현 조절 과정

1. 전사 조절
염색질 구조의 응축 정도가 약할 때 RNA 중합 효소 등의 전사
기구가 DNA에 잘 결합할 수 있다.
◇염색질 구조
진핵 세포의 DNA는 히스톤 단백질에 감겨서 뉴클레오솜이라는
기본 단위를 형성한다. 뉴클레오솜은 더 응축되어 염색질 구조
를 가진다. 응축된 염색질은 RNA 중합 효소가 프로모터에 결합
할 수 없어 전사가 불가능하다.
프로모터 앞쪽의 조절 요소에 전사 조절 인자가 결합하여 유전자 발현을 조절한다. 전사 조절 인자 중 전사를
촉진하는 것을 전사 촉진 인자, 억제하는 것을 전사 억제 인자라고 한다. → 전사 촉진
인자는 염색질의 구조를 풀어 주거나 전사기구의 형성을 촉진함으로써 전사 개시가
잘 일어나도록 한다.
◇ 진핵생물의 전사
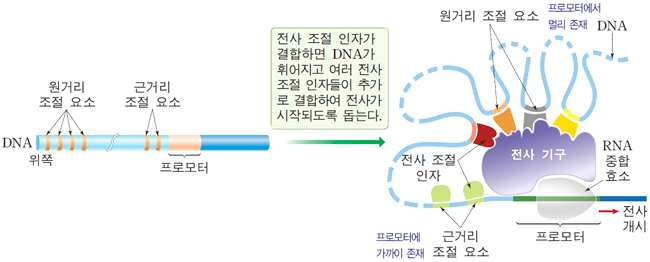
원핵생물과 비교하여 큰 차이점은 진핵생물의 전사 과정에는 다양한 전사 조절 인자들이
결합하여 전사를 조절한다는 데 있다. 따라서 진핵생물에서 프로모터는 원핵생물에 비
해 아주 복잡하게 구성되어 있다.
◇ 전사 개시의 과정
전사 조절 인자 중 전사 촉진 인자가 조절 요소에 결합하면 DNA를 꺾어 주는 단백질에 의해 DNA가 꺾이고 이로 인해 전사 촉진 인자들이 프로모터 가까이에 위치하게 된다. RNA 중합 효소, 매개 단백질, 전사 조절 인자 등이 결합하여 복합체인 전사 기구가 형성되면서 전사가 시작된다.
2. 전사 후 조절(RNA 가공 조절)

mRNA 말단의 변화 : 전사 직후 형성된 mRNA 전구체의 5´ 말단에 캡이 형
성되며, 3´말단에 폴리A 꼬리가 형성된다.
※ 5´캡과 폴리 A 꼬리 : mRNA의 가수 분해를 억제하고, 핵에서 세포질로의
수송, 리보솜과 mRNA의 결합 촉진 등에 관여한다.
RNA 스플라이싱 : 전사 직후의 mRNA는 엑손과 인트론이 모두 존재하여 단
백질 합성에 바로 이용될 수 없어 인트론이 제거되고 엑손끼리 서로 연결
되는 스플라이싱이 일어난다. 엑손의 조합에 따라 다양한 종류의 mRNA가 생성된다.
3. RNA 수송 조절
성숙한 mRNA가 핵을 통해 세포질로 배출되는 속도가 조절된다.
4. RNA 분해 조절
mRNA는 폴리 A 꼬리가 짧아지면서 분해되는데, mRNA의 분해 속도를 조절하여 합성되는 단백질의 양을 조절한다.
5. 번역 조절
단백질 합성의 개시를 조절하는 단백질에 의해 단백질 번역을 조절할 수 있다.
6. 번역 후 조절(단백질 변형 과정상의 조절)
진핵 세포의 단백질은 합성된 후 기능적인 단백질로 변형되는 과정을 거치는데, 단백질의 화학적 변형이나 기능 조절, 수송 과정 등이 조절될 수 있다.
단백질이 세포 속에서 얼마 동안 역할을 할 것인가는 단백질의 분해 속도를 통해 조절한다.
(요약)
핵산 대사
- 유전자 구조
①구조유전자 :
원핵생물의 구조유전자는 모두 단백질 또는 RNA에 해당하는 정보이나
진핵생물은 단백질 또는 RNA에 대응하는 엑손과 대응하지 않는 인트론으로 구성
②조절유전자
개시코돈 : AUG, GUG(드물게)
종결코돈(=넌센스코돈) : UAA, UGA, UAG
DNA복제과정 : 개시, 신장, 종결
신장
①선도가닥 : 3′→5′ 주형
②지연가닥 : 5′→3′ 주형, 오카자키단편
DNA복제 수복 : 틀린 상보적 염기 결합수복, 염기제거수복, 디옥시리보뉴클레오티드 절제 수복, 직접수복
전사 : RNA가 합성되는 과정
전사 개시
①대장균(세균) : 촉진유전자(promoter)-프리브노 상자
②진핵생물 : 촉진유전자- TATA상자와 Inr서열
③DNA와 달리 시발물질이 필요하지 않음
전사 주형사슬 : 3′→5′
cDNA : 상보적 DNA, 시발물질 필요, 오류빈도 매우 높음→변원성 바이러스가 빈번하게 발생하는 원인
mRNA
①진핵세포 : 스플라이싱, 5′cap, 3′말단의 폴리 A 꼬리(분해방지)
②원핵세포 : 3′말단의 폴리 A 꼬리(분해 촉진)
rRNA
①진핵세포 : 45s전사
②원핵생물 : 30s전사, tRNA포함
번역 : mRNA의 유전정보에 근거하여 단백질이 합성되는 것.
폴리솜 : mRNA상의 여러 개의 리보솜이 결합→연속적으로 단백질 합성
대장균(세균)의 개시코돈에 대응하는 아미노산이 포르밀메티오닌인 것은 샤인-달가르노 배열 때문
진핵세포의 개시코돈은 메티오닌이 운반된다.(단, 미토콘드리아와 엽록체에서 단백질이 합성될 때는 포르밀메티오닌인 운반됨)
폴리펩티드상 아미노산이 정확한 순서로 결하는 이유 : tRNA의 안티코돈이 mRNA의 코돈에 적확하게 결합
단백질 구조 완성 : 입체구조, 아미노기말단의 포르밀메티오닌, 신호배열, 인산화, 오리고당의 결합
하우스키퍼 유전자 : 세포에 필수적인 유전정보
전사인자 :
①특이성 인자 : 프로모터를 인식하는 특성을 변화시켜.
②억제자 : 억제, 부조절(네거티브)
③활성자 : 촉진, 양조절(포지티브)
오퍼레이터 : 억제자가 결합하는 DNA상의 부위
증강자(엔헨서) : 활성자가 결합하는 DNA상의 부위
오페론 : 세균에서 유전자의 발현을 동시에 조절하는 영역
(요약2)
⦁어미세포 DNA의 구조에 근거하여 복제되며, 복제과정 중에 오류가 있는 경우 이를 수정하는 체계가 있기 때문이다. 복제오류를 수
정하는 방법은 복제과정 중에 틀린 상보적 염기의 deoxyribonucleotide가 주형 polydeoxyribonucleotide에 결합된 것을 제거하
고 옳은 deoxyribonucleotide를 결합, 화학변화에 의해 손상된 염기를 가지는 deoxyribonucleotide 혹은
oligodeoxyribonuclotide 단편을 제거하고 옳은 deoxyribonucleotide 혹은 oligodeoxyribonucleotide 단편으로 교환 결합, 손
상된 염기의 수복 등이다.
⦁원핵세포의 경우는 RNA 중합효소가 DNA의 mRNA의 전사가 시작될 5′쪽 -10 및 -35 염기쌍 전방에 존재하는 promoter를 인식
하여 결합하면 RNA의 전사가 시작된다. 진핵세포의 경우는 RNA 중합효소Ⅱ가 DNA의 mRNA의 전사가 시작될 5′쪽 -30 및
+1bp에 존재하는 promoter의 공통구조에 가까운 구조를 가지는 promoter는 RNA중합효소가 잘 결합하기 때문에 mRNA 합성이
활발하여 많이 발현된다.
⦁생물체에서 필수적으로 요구되는 Housekeeping gene의 발현 정도는 이 유전자의 발현을 위한 mRNA 합성 개시 promoter의
구조와 RNA 중합효소의 친화성에 따라 결정된다. promoter의 공통구조에 가까운 구조를 가지는 promoter는 RNA 중합효소가
잘 결합하기 때문에 mRNA 합성이 활발하여 많이 발현된다.
⦁bacteriophage는 크기가 4만~5만bp인 것만이 단백질로 포장되어 숙주세포로 감염될 수 있기 때문이다.
⦁cosmid는 bacteriophage의 감염 형태로 숙주세포로 들어가야 한다. cos서열은 단백질 표피에 의한 포장 및 숙주세포에 감염되는
데 필요한 유전정보를 가지고 있어 cosmid가 숙주세포로 bacteriophage와 동일한 방법으로 감염될 수 있도록 한다.
⦁원핵세포의 구조유전자는 모두 단백질 혹은 RNA에 해당하는 정보이나, 진핵세포 구조유전자의 경우는 단백질 혹은 RNA에 대응하
는 엑손(exon)과 대응하지 않아 번역되지 않는 인트론(intron) 부위로 구성되어 있다.
⦁유전암호란 mRNA상의 1개의 아미노산에 대한 3개의 ribonucleotide의 5′→3′방향 배열이다. 단백질의 합성을 개시하는 유전기호
는 AUG이다.
⦁진핵세포 전사의 중추 역할을 담당하는 RNA 중합효소Ⅱ의 촉진유전자는 -30 염기쌍 부위, +1 염기쌍 부위의 공통적인 염기쌍 배
열을 가지는 곳과 공통성이 없는 조절부위가 존재한다. -30염기쌍 부위와 +1염기쌍 부위의 공통구조는 각기 5′TATAAAA3′. 5′
YYANTAYY3′이며 각각 TATA 상자, Inr 서열(Initiator, 개시자)이라한다. 원핵세포의 촉진유전자는 프리브노(Pribnow)상자이다.
⦁원핵생물에서 개시코돈인 5′AUG가 polypeptide 첫 번째 아미노산인 N-formylmethionine에 해당하는 유전암호로 인식되는가 혹
은 polypeptid 중간의 methionine에 해당하는 유전암호로 인식되는가는 샤인-달가르노 배열이 mRNA상의 AUG 유전기호로부터
5′방향으로 8~13 염기 부근에 존재하는가 여부에 의해 결정된다.
⦁진핵생물에서 전사에 의해서 합성된 mRNA는 단백질 구조로 번역되니 않는 intron 부위는 splicing에 의해 제거되기 때문에 완성
된 mRNA를 역전사하여 필요한 단백질의 유전자를 합성할 수 있다.
E 연습문제
1. DNA의 복제가 매우 정확하게 이루어져 어미세포와 딸세포가 동일한 유전정보를 가질 수 있는 근거는 무엇인가?
→ 어미세포 DNA의 구조에 근거하여 복제되며, 복제과정 중에 오류가 있는 경우 이를 수정하는 체계가 있기 때문이다. 복제오류를 수정
하는 방법은 복제과정 중에 틀린 상보적 염기의 deoxyribonucleotide가 주형 polydeoxyribonucleotide에 결합된 것을 제거하고 옳
은 deoxyribonucleotide를 결합, 화학변화에 의해 손상된 염기를 가지는 deoxyribonucleotide 혹은 oligodeoxyribonucleotide 단편
을 제거하고 옳은 deoxyribonucleotide 혹은 oligodeoxyribonucleotide 단편으로 교환 결합, 손상된염기의 수복 등이다.
2. DNA 구조에 근거하여 mRNA 전사가 시작되는 과정을 제시하시오.
→ 원핵세포의 경우는 RNA 중합효소가 DNA의 mRNA의 전사가 시작될 5′쪽 -10 및-35 염기쌍 전방에 존재하는 promoter를 인식하여
결합하면 RNA의 전사가 시작된다. 진핵세포의 경우는 RNA 중합효소Ⅱ가 DNA의 mRNA의 전사가 시작될 5′쪽 -30및 +1bp에 존재
하는 promoter를 인식하여 결합하면 RNA의 전사가 시작된다.
3. 생물체에서 필수적으로 요구되는 형질이 발현되는 정도가 조절되는 방법은 무엇인가?
→ 생물체에서 필수적으로 요구되는 Housekeeping gene의 발현 정도는 이 유전자의 발현을 위한 mRNA 합성 개시 promoter의 구조
와 RNA 중합효소의 친화성에 따라 결정된다. promoter의 공통구조에 가까운 구조를 가지는 promoter는 RNA 중합효소가 잘 결합하
기 때문에 mRNA 합성이 활발하여 많이 발현된다.
4. bacteriophage가 큰 유전자만 클로닝할 수 있는 이유는 무엇인가?
→ bacteriophage는 크기가 4만∼5만bp인 것만이 단백질로 포장되어 숙주세포로 감염될 수 있기 때문이다.
5. cosmid의 cos 서열의 중요성은 무엇인가?
→ cosmid는 bacteriophage의 감염 형태로 숙주세포로 들어가야 한다. cos 서열은 단백질 표피에 의한 포장 및 숙주세포에 감염되는
데 필요한 유전정보를 가지고 있어 cosmid가 숙주세포로 bacteriophage와 동일한 방법으로 감염될 수 있도록 한다.
기본교재 연습문제 해설
1. 엑손과 관련된 내용은?
① 원핵세포의 유전자 발현을 조절한다. ② 진핵세포의 아미노산 혹은 RNA의 구조에 대응하는 유전정보이다.
③ 진핵세포의 단백질 구조로 발현되지 않는 DNA의 부위이다. ④ 원핵세포의 RNA에 해당하는 DNA 구조이다.
정답 ② 원핵세포의 구조유전자는 모두 단백질 혹은 RNA에 해당하는 정보이나, 진핵세포 구조유전 자의 경우는 단백질 혹은 RNA에 대
응하는 엑손(exon)과 대응하지 않아 번역되지 않는 인트론(intron) 부위로 구성되어 있다.
2. 단백질 합성의 개시를 지시하는 유전암호는?
① DNA에 존재하는 TAA ② DNA에 존재하는 UAG ③ mRNA에 존재하는 UGA ④ mRNA에 존재하는 AUG
정답 ④ 유전암호란 mRNA상의 1개의 아미노산에 대한 3개의 ribonucleotide의 5′→3′방향 배열이다. 단백질의 합성을 개시하는 유전기
호는 AUG이다.
3. 진핵세포의 촉진유전자에 해당하는 것은?
① 프리브노 상자, TATA 상자 ② TATA 상자, Inr 서열 ③ Inr 서열, 프리브노 상자 ④ 프리브노 상자, 오페론
정답 ② 진핵세포 전사의 중추 역할을 담당하는 RNA 중합효소Ⅱ의 촉진유전자는 -30 염기쌍 부위,+1 염기쌍 부위의 공통적인 염기쌍
배열을 가지는 곳과 공통성이 없는 조절 부위가 존재한다. -30 염기쌍 부위와 +1 염기쌍 부위의 공통구조는 각기 5′TATAAAA3′, 5′
YYAN T YY3′이며 각각 TATA 상자, Inr 서열(Initiator, 개시자)이라 한다. A원핵세포의 촉진유전자는 프리브노(Pribnow) 상자이다.
4. 원핵생물 단백질의 첫 번째 아미노산이 methionine이 아니고 N-formylmethionine 인 원인에 해당하는 것은?
① 샤인-달가르노 배열 ② 개시코돈의 변이 ③ 특이성 인자 ④ 자율복제배열
정답 ① 해설 원핵생물에서 개시코돈인 5′AUG가 polypeptide 첫 번째 아미노산인 N-formylmethionine에 해당하는 유전암호로 인식되
는가 혹은 polypeptide 중간의 methionine에 해당하는 유전암호로 인식되는가는 샤인-달가르노 배열이 mRNA상의 AUG 유전기호로
부터 5′방향으로 8~13 염기 부근에 존재하는가 여부에 의해 결정된다.
5. 진핵생물의 유전자를 분리할 때 mRNA를 이용하는 것이 편리한 점은 무엇인가?
① mRNA는 직접 게놈 도서관으로 이용할 수 있다. ② mRNA를 PCR의 주형으로 사용하여 PCR을 실시하여 유전자를 간단하게 분리
할 수 있다. ③ mRNA는 DNA의 인트론 부위의 상보적인 염기의 ribonucleotide로 구성되어있다.
④ mRNA는 DNA 엑손의 상보적인 염기의 ribonucleotide로 구성되어 있다.
정답 ④ 진핵생물에서 전사에 의해서 합성된 mRNA는 단백질 구조로 번역되지 않는 intron 부위는 splicing에 의해 제거되기 때문에 완성
된 mRNA는 단백질로 번역되는 엑손 부위로만 구성되어 있다. 그러므로 mRNA를 역전사하여 필요한 단백질의 유전자를 합성할 수 있다.
[인강연습문제]
1. 촉진유전자의 기능은 무엇인가?
☞전사되어야 하는 유전자의 앞쪽에 특별한 구조를 가지고 있는 부위로서 RNA중합효소가 결합하면 DNA가 풀리면서 mRNA가 합성되
기 시작하도록 하는 조절 유전자 부위이다.
2.원핵세포 단백질의 첫번째 아미노산인 formyl metionine이 분해되어 없어지는 경우는 어느 때인가?
☞ 원핵세포 단백질의 첫번째 아미노산인 formyl metionine이 분해되어 없어지는 경우는 어느 때인가?
※① 두번째 아미노산이 alanine, serine, threonine 이면 methionyl aminopeptidase에 의해 formyl metionine이 분해되어 없어질
수 있다.
② N-말단에 신호배열이 존재하는 경우에는 합성된 polypeptide가 목표로 하는 장소에 운송된 후에 신호배열은 분해, 제거된다.
3.필수 유전자(구성적 유전자)의 발현과 비필수 유전자의 발현이 조절되는 차이는 무엇인가? ☞ 필수 유전자는 항상 발현되면서 발현
되는 량만 조절된다. 비필수 유전자는 필요에 따라 발현되거나 발현되지 않는다.
4.박테리오파지 DNA가 유전자 운반체로서의 장점은 무엇인가?
☞10,000~23,000bp의 큰 유전자를 운반할 수 있다.
5. 진핵세포의 유전자를 분리할 때는 mRNA를 사용하는 것이 유리한 이유는?
※mRNA는 유전자에 근거하여 합성된 후에 인트론에 해당하는 부위는 splicing에 의해 분해되고 단백질에 해당하는 엑손부위만이 존재
한다. 그러므로 mRNA를 역전사하여 DNA를 합성하여 이용하면 형질을 발현시켜 해당 물질을 생산할 수 있다.
6. 단백질 합성의 개시를 지시하는 유전암호는 무엇인가?
1. DNA에 존재하는 TAA 2. mRNA에 존재하는 UGA
3. DNA에 존재하는 UAG 4. mRNA에 존재하는 AUG
※ 부족하지만 글의 내용이 도움이 조금이라도 되셨다면, 단 1초만 부탁드려도 될까요? 로그인이 필요없는 하트♥(공감) 눌러서 블로그 운영에 힘을 부탁드립니다. 그럼 오늘도 행복한 하루 되십시오^^
'농업생물화학' 카테고리의 다른 글
| 농업직, 농학과, 화학, 방통대 등 농업생물화학 핵심 요점 정리 15. 가축의 생산 (4) | 2022.12.05 |
|---|---|
| 농업직, 농학과, 화학, 방통대 등 농업생물화학 핵심 요점 정리 14. 작물의 생산 (14) | 2022.12.04 |
| 농업직, 농학과, 화학, 방통대 등 농업생물화학 핵심 요점 정리 12. 단백질 대사 (3) | 2022.12.02 |
| 농업직, 농학과, 화학, 방통대 등 농업생물화학 핵심 요점 정리 11. 지방질 대사 (10) | 2022.12.01 |
| 농업직, 농학과, 화학, 방통대 등 농업생물화학 핵심 요점 정리 10. 탄수화물 대사 (5) | 2022.11.30 |