16. 핵산의 복제와 유전자의 전사 및 발현
들어가기
⋅원핵세포들은 핵이 없이 DNA가 세포질에 노출되어 있고, 세대 기간이 짧아서 모든 일들을 거
의 동시에 수행해야 함
⋅진핵세포들은 핵에서 전사된 RNA 분자들이 세포질로 나와야 단백질이 합성될 수 있음
⋅진핵세포의 유전물질 발현 및 조절과정은 원핵세포보다 더 복잡함
1. 유전자 구조
⋅세균이나 바이러스에서 사람에 이르기까지 모든 생물체들은 각자의 유전체(genome) 분자를 가짐
※유전체 : 한 생명체의 세포가 갖고 있는 총 유전정보를 칭함. 이를 지놈이라 읽음
※염색체 : 생명체 세포가 갖는 유전물질 DNA는 1개 이상의 분자로 이루어져 있는데, 이렇게 기
다란 크기의 DNA는 히스톤 단백질 덩어리를 둘러싸고 포장되어 세포주기에 따라 그 구조가
달라지며 세포분열 시기에는 아주 밀집된 형태로 뚜렷한 형태를 갖는 특징적 개수로 나타남
1) 유전자의 구성
⋅유전자(gene)은 전사되어 하나의 폴리펩티드 또는 일차 RNA(hnRNA, pre-tRNA, pre-rRNA)
을 만들어 낼 수 있는 길이의 DNA 부분
※유전자 : 하나의 폴리펩티드 또는 RNA 분자를 만들어 내는 DNA 부위 전체를 말함
⋅원핵세포의 대부분 유전자 → mRNA 한 개에서 여러 개의 단백질을 만듦
⋅진핵세포의 대부분 유전자 → mRNA 한 개에서 단백질 한 개를 만들어 냄
⋅진핵세포의 유전자는 엑손(exon)과 인트론(intron)으로 나뉨
※엑손 : 단백질의 아미노산 서열을 지정하는 mRNA 부분을 구성
※인트론 : 1차 전사체로는 전사되지만 세포질에 나온 mRNA에는 나타나지 않는 부분
⋅헤모글로빈의 β-단백질 유전자는 인트론 하나가 유전자의 반 이상을 차지함
(예) 오브알부민 유전자 → 여러 개의 짧은 크기의 엑손들이 흩어져 있음
⋅반면에 히스톤 유전자는 인트론이 없이 엑손 하나를 가짐
⋅인트론 기능은 아직 정확히 알려져 있지 않음
2) 염색체상의 유전자
⋅인간의 46개 염색체상에는 약 31억 쌍의 염기쌍이 있음 → 적어도 29,000여개의 유전자를 갖
는다고 알려져 있음
⋅실제로 생물학적 기능을 보이는 유전자는 훨씬 많음
⋅그러므로 유전자 숫자는 중요하지 않으며, DNA의 길이가 길다가 더 많은 유전적 정보를 가진
것이라고 볼 수 없음
3) 원핵세포의 염색체 DNA의 특징
⋅전형적인 원핵생물의 유전체는 진핵세포와 기본 DNA 구조는 같지만 특징적 차이가 있음
-하나의 커다란 원형 DNA 분자로 되어 있음
-유전체 DNA 이외에도 아주 작은 크기의 플라스미드 DNA를 갖는 경우도 있음
-DNA 분자들은 초나선형 구조로 상당히 꼬여 있고, 특히 유전체 DNA는 루프 구조로 뭉쳐
있으며, 주로 세포질에 떠 있는 ‘핵양체(nucleoid)'로 되어 있음
-포장 해결방법으로는 DNA를 초나선형 감김(super coil) 상태로 존재하게 해야 함
-초나선형 감김은 ‘감긴(colied)' DNA가 또 감긴(coiling) 것을 표현한 것임
| ※ 복제 : 이중나선 DNA의 두 가닥이 분리되고 주형가닥으로 사용되어 새로운 뉴클레오티드들이 정확히 상보적 염기쌍으로 복사함으로써 새로운 두 쌍의 유전체를 만드는 과정 ※ 전사 : 두 가닥의 DNA 사슬 중 한 쪽 가닥의 유전정보가 RNA 유전정보로 전환되는 과정 ※ 단백질 합성 : 전사된 mRNA의 유전정보를 토대로 아미노산 순서가 결정되어 펩티드결합으로 폴리펩티드를 합성하는 과정 ※ 역전사 : RNA 분자 염기서열이 DNA 분자 염기서열로 전환되는 과정 |
4) 진핵세포의 염색체 DNA 특징
⋅진핵세포들의 DNA는 핵 속에 있음
⋅세포분열이 끝난 세포의 DNA들은 히스톤 단백질 옥타머(8개 단백질)를 포장하듯이 감싸서
뉴클레오좀 덩어리를 이룸
⋅핵막은 핵공(nuclear pore)을 갖고 있어서 핵 내외로 분자들을 통과시킴
⋅히스톤, DNA 복제효소, RNA 중합효소 등이 세포질에서 핵으로 들어가고, 핵에서 전사된 여러
크기의 RNA 분자들은 세포질로 나옴
(1) 진핵세포의 염색체 DNA
⋅대부분 여러 DNA로 나뉘어져 있고, 서로 다른 크기의 대단히 긴 분자
⋅염색약에 염색되므로 염색체 DNA라고도 부름
⋅원핵세포는 대부분 1개 염색체를 갖지만, 진핵세포는 암컷과 수컷에서 하나씩을 받아서 두 개
의 상동염색체로 존재함
(2) 세포주기에 따른 DNA의 특징
⋅진핵세포들은 세포주기에 따라 세포가 분열하고 성장함
-체세포분열이 끝난 세포는 GI 단계로 들어감 → 핵막이 다시 생김 → 염색체 DNA는 덜 촘촘
한 진정염색질로 바뀜 → 유전자 전사활동이 활발함
-세포주기 S 단계에 오면 DNA들이 복제
-세포주기 G2 단계에 오면 복제가 끝난 DNA들이 히스톤 단백질에 포장된 염색분체
(chromatid)상태로 체세포 분열 단계로 들어감
-체세포 분열단계(M)에서 염색체들은 분리되어 두 개의 G1 단계세포로 나뉨
⋅진핵세포들은 세포주기에 따라 세포가 분열하고 성장함
분열된 세포는 촘촘한 이질 염색질 구조 & 덜 촘촘한 진정 염색질 & 완전히 DNA가 노출된
부분 등으로 시간에 따라 역동적으로 바뀜
이러한 DNA 상태를 염색질(chromatin)이라 부름
염색질은 핵에 분산되어 있어서 잘 보이지 않지만, 분열시에는 촘촘한 뉴클레오좀들이 염색체
로 바뀜
2. DNA 복제
1) DNA 복제 원리
(1) 반보존적 DNA 복제
⋅‘DNA 이중나선 모델’이 발표된 이후, 복제기작에 대해 반보존적, 보존적, 무작위 분산적 가설이
각각 제시됨
⋅메셀슨 & 스탈 → DNA 복제는 반보존적이여서 복제 전의 한 가닥이 새로이 합성된 가닥과
염기쌍을 이룬 다고 알려짐
| ※ 세포 주기 : 진핵세포는 한 세포분열에서 다음 세포분열까지 4단계의 세포주기를 갖는데, G1 단계는 세 포대사과정이 활성화되고 있는 단계이며, 다음 S단계는 DNA가 복제되는 단계이고, G2 세포분열 준비단 계를 지나 곧 체세포분열(M)단계로 염색체와 세포질이 나누어짐 -분열된 세포는 촘촘한 이질 염색질 구조 & 덜 촘촘한 진정 염색질 & 완전한 DNA가 노출된 부분 등으로 시간에 따라 역동적으로 바뀜 -이러한 DNA 상태를 염색질(chromatin)이라 부름 -염색질은 핵에 분산되어 있어서 잘 보이진 않지만, 분열 시에는 촘촘한 뉴클레오좀들이 염색체로 바뀜 ※ 뉴클레오좀 : 염색질의 기본 반복단위로서 4가지 히스톤 단백질이 두 개씩 모여서 8개의 옥타머 (octamer)가 되어 약 146개 염기쌍의 DNA 길이가 그룰 둘러싸면서 포장된 단위를 말함 ※ 염색질 : 체세포 분열 후 핵 내에 존재하는 느슨하게 풀어진 상태의 염색체 구조로서 현미경상에서 체세 포분열 때의 염색체와 다른 구조를 보여줌 |
(2) 복제시작점과 양방향성 복제
⋅케언즈(Carins) → 한 곳의 복제시작점을 알아냄
⋅DNA의 AT 염기쌍이 풍부한 지역을 선택적으로 변성시켜 복제시켰더니 양쪽으로 버블
(bubble)이 커져가는 것을 확인함
⋅즉, 두 DNA 가닥이 동시에 복제되고, 원형의 세곤 염색체 복제는 양쪽 방향으로 진행됨
⋅원형의 DNA 분자는 하나의 복제시작점(ori site)에서 양쪽 방향으로 복제됨
⋅진핵세포 DNA와 같이 일직선의 긴 DNA들은 시작점이 여러 개 있으며 각 시작점에서 양쪽방
향으로 복제를 시작함
2) 원핵세포 DNA의 복제
⋅세균의 DNA 복제 : 복제시작점에서 시작 → 나선형 DNA 부분이
더욱 풀리면서 복제 → 신장과정 → 복제 완성 → 종결
(1) 개시단계 : 복제시작점과 DNA 나선의 풀림
⋅복제 개시단백질(DnaA) + 복제시작점과 결합 → 약간의 DNA 부분이 풀림 → 헬리케이즈
(helicase)효소가 수소 결합을 끊음 → 나선형을 조금 더 길게 풀어줌
⋅단일가닥결합(SSB) : 단백질이 풀어진 단일가닥 DNA에 결합하여 복제되는 동안 풀린 상태를
일정기간 유지하게 함
⋅DNA 자이레이즈(gyrase) : 회전압력을 감소시켜 나선형을 풀리게 함
⋅DNA 복제효소 III : 새로운 뉴클레오티드를 주형 DNA 가닥의 염기들과 상보적으로 염기쌍이
이루어지도록 함
⋅프리메이즈(primase) 효소 : 일부 DNA 가닥을 주형으로 삼아 약 10~12개 길이의 RNA 프라
이머를 합성
(2) 신장단계
⋅RNA 프라이머가 만들어지면 주형 DNA 가닥의 염기와 염기쌍을 이루는 새로운 DNA 뉴클레
오티드가 프라이머의 3‘-OH에 연결되면서 DNA 복제가 계속되고 새로운 가닥이 신장되어 감
⋅DNA 복제효소 I &II :DNA 합성
⋅DNA 복제효소 III(DNA pol III)은 DNA를 복제하는 효소로서 10개의 단위 단백질체로 구성
⋅신장되고 있는 DNA 가닥의 3‘-말단에 새로운 뉴클레오티드를 첨가시켜 5’-말단에서 3‘-말단으
로 DNA가 신 속히 길어지게 함
⋅이 때, 잘못 들어온 염기의 뉴클레오티드가 들어오면 DNA 복제효소 III의 복합체에서 핵산말
단가수분해효소 활성을 가진 효소가 제거함
⋅1960년대 오카자키(Okazaki)가 DNA 한쪽 가닥이 짧게 합성된 후, 이것들이 다시 이어지는
것을 발견함
⋅RNA 프라이머와 연결되어 있는 약 1,000개 길이의 짧은 단편을 ‘오카자키 조각’이라 부름
→ 한 쪽은 첫 번째 프라이머에서부터 연속적으로 합성되고(선도가닥, leading strand),
→ 다른 쪽은 불연속적으로 합성되는 것(지연가닥, lagging strand)
| ※ 복제 시작점 : 대장균 같은 우너핵세포의 DNA 복제 시작은 한 군데에서 양쪽 방향으로 복제가 시작됨 ※복제분기점 : 복제가 시작하면 두 가닥 DNA가 풀려나가면서 Y자 모양의 분기점이 복제 방향으로 나아감 ※주형 : 핵산의 상보적 가닥 합성에 필요한 정확한 정보를 보여 주는 DNA 또는 RNA 사슬. RNA 합성은 DNA 주형이 필요하고, DNA 복제에는 상보적인 양쪽 DNA 사슬이 주형으로 쓰임 ※프라이머 : DNA 주형에 있는 짧은 길이의 RNA로서 복제개시점에서 DNA 뉴클레오티드가 부착할 수 있 도록 3‘-OH기를 제공함. 짧은 RNA 조각이 프리마제(primase)효소에 의해 일시적으로 만들어진 것 |
(3) 종결단계
⋅두 가닥의 복제분기점이 서로 반대 방향에서 돌아오면서 만나면 복제가 종결됨
⋅앞쪽의 뉴클레오티드와 인산디에스테르결합이 끊어짐 틈(nick)을 DNA 연결효소가 연결시켜
DNA 복제를 사실상 완결시킴
(4) 복제의 정확성
⋅정확한 복제가 가능한 이유는 교정(proofreading) & 수선(repair) 때문
-교정 : 핵산말단가수분해효소 활성
-수선 : 잘못 연결된 부정한 DNA는 또 다른 복제효소 분자가 잘못된 염기짝을 제거하고 교정시킴
3) 진핵세포의 DNA 복제
⋅원핵세포의 DNA 보다 길고 복잡한 구조를 형성하고 있음
-여러 개의 복제 시작점(ori site)을 가짐
-선형의 DNA를 가지므로 끝부분의 말단소체9telomere)구조의 복제 시작에는 텔로머레이즈
(telomerase)효소가 작용함
-뉴클레오좀 구조를 가지므로 감겨 있는 DNA를 풀어주어야 함
(1) 복제시작점
⋅여러 곳에 있으며, AT 염기쌍 > GC 염기쌍
⋅세포주기 동안 단 한번만의 복제가 이루어지도록 조절
⋅두 단계의 복제시작점을 가짐 -첫 번째 : 복제가 일어나도록 준비하는 단계 -두 번째 : 복제가
준비된 복제시작점에서만 복제가 시작
(2) 이중나선 DNA의 풀림
⋅단백질들이 DNA를 풀어주고 단일가닥결합(SSB) 단백질들이 이를 유지시켜 주어 복제가 시작되도록 함
(3) 진핵세포 DNA 복제 효소
⋅복제효소 → 복제, 재조합, DNA 수선 등에 관여
⋅진핵세포는 5가지 서로 다른 복제효소 α, β, γ, δ, ε 등을 가짐
-복제효소 α : 여러 단위체 단백질들의 복합체, 프리메이즈. 복제 활성 있음 & 핵산말단가수분
해효소 없음
-복제효소 δ : 원핵세포의 DNA pol III과도 같은 역할을 함. 오카자키 조각에 있는 RNA 프
라이머는 RNA 분해효소(RnaseH)가 제거시킴
-복제효소 β, ε : DNA 복구과정에 관여함
-복제효소 γ : 미토콘드리아 DNA를 복제하는데 쓰임
⋅진핵세포 DNA 복제기구와 원핵세포와의 차이점
-유전체의 크기가 엄청 큼
-원핵세포는 염색체가 하나이지만, 진핵세포는 염색체 46개 모두 복제해야 함
-원핵세포 염색체는 원형이지만, 진핵세포는 일직선의 선형임. 선형의 DNA는 매번 복제 때마
다 끝 부분이 조금씩 없어짐
| ※ 틈 : 뉴클레오티드와 뉴클레오티드가 연결될 때 인산다이에스테르 결합이 안 되어 서로 끊어져 있는 상태를 말함. DNA 리가아제(ligase, 연결효소)가 이를 연결해줌 |
3. DNA 변이와 수선
⋅환경으로부터 방사선, 화학적인 돌연변이원, 자연발생적인 변화 등에 노출되어 손상을 입게 됨
⋅자주 일어나지는 않지만, 손상이 치명적일 수 있고, 그로 인한 변이가 세대에서 세대로 이어질 수도 있음
1) DNA 돌연변이와 돌연변이원
⋅DNA 복제과정 중 실수로 서로 맞지 않는 염기쌍 결합이 일어나면 돌연변이(mutation)을 일으킴
⋅돌연변이가 일어난 유전자는 아미노산 서열이 바뀐 단백질을 생산하여 세포의 정상적인 기능을 손상시킴
⋅여러 효소 및 단백질들이 관여하는 복구시스템을 작동시켜 원래 DNA로 복구시킴
2) 돌연변이원에 의한 DNA 손상
(1) 염기의 알킬화
⋅DNA에서 음전하를 띠는 인산과 부분적인 음전하를 띠는 염기에 주변의 전자 친화물들이 공격
하여 알킬기를 추가시켜 알킬화 시킴
⋅알킬화된 염기들 → 복제 시 실수를 일으킴 → 돌연변이가 될 확률 ↑
(2) 자외선
⋅DNA 상의 연속된 피리미딘 염기를 연결시켜 피리미딘 이합체(pyrimidine dimers)를 만듦
⋅염기쌍 결합을 깨트려서 복제가 중단 or 다음 세대에 부정확한 염기쌍이 형성
(3) 감마선 및 X-선
⋅에너지가 큰 감마선과 X-선은 DNA 주변의 물 분자를 이온화 시킴 → DNA에 손상을 가함
⋅산소 or 질소 자유라디칼을 만들어 → DNA를 공격함 → DNA 가닥이 끊어지기도 함
3) 일반적인 DNA 복구과정
⋅DNA 복구시스템은 대부분의 생물체 내에 존재함
⋅대개 변경되지 않은 주형가닥의 염기 순서를 이용하여 변경된 가닥의 염기들을 아래와 같이복구함
① 변경된 염기 인식
② 그 염기를 제거
③ 그 틈을 DNA 복제효소와 DNA 연결효소가 복구
. DNA 수선 기작에는 뉴클레오티드 두 가닥 모두 필요함
. 수선 교정 기작에는 4 종류가 있음
-부정한 DNA 염기쌍의 수선
-직접수선
-염기절제 수선
-뉴클레오 절제 수선
(1) 부정합 DNA 염기쌍의 수선
⋅부정합 염기짝의 복구능력은 복제효소가 갖고 있음
⋅이 작용은 염기쌍 간의 수소결합이 잘못되면 복제효소의 합성속도가 느려짐 → 효소의 합성작
용 활성부위에 서 핵산말단가수분해효소 활성부위로 효소 작용이 바뀌게 됨
⋅DNA pol I(부정함 DNA 수선 효소) : 잘못 들어온 염기쌍의 비틀어진 부분을 잘라내고, 본래
의 DNA 가닥을 주형으로 내세워 새로운 뉴클레오티드로 빈 공간을 채워줌
| ※ 말단소체 : 진핵세포 염색체 DNA의 말단 부분 ※ 텔로머레이즈 : 단백질과 RNA로 구성된 효소로, 말단소체를 복세키니다. 효소의 RNA는 말단소체의 반복서열에 상보적이다 |
(2) 직접수선
⋅손상된 지역을 직접 제거한 후 복구하는 경우
⋅빛에 의해 재활성화되는 DNA 광분해효소 이용 → 피리미딘 이합체를 직접 분해하여 정상적인
피리미딘으로 북구해냄
(3) 염기절제수선(base excision repair)
⋅손상된 염기들은 DNA 글리코실 가수분해효소로 끊음 → AP(apurinic) 핵산내부 가수분해효소
가 자름 → 디옥시리보오스 인산디에스테라아제 효소가 인산 단위로 자른 후 → 다시 DNA
pol I이 복구함
(4) 뉴클레오티드 절제 수선
⋅피리미딘 이합체처럼 부피가 커진 DNA 손상을 제거하는 과정
⋅가장 중요한 수선과정의 하나임
⋅관련되는 효소 복합체가 구조적으로 비틀려있는 지역을 찾아내어, 두 가닥을 서로 분리하여 단
일가닥결합 (SSB) 단백질을 결합시켜 안정화시킴
⋅손상된 양 가닥의 당-인산 골격 절단 제거된 부분은 DNA pol I이 메우고 DNA 연결효소가
봉합함
4. 재조합
⋅재조합은 DNA 분자 간의 물리적 교환
⋅교환이 유사한 DNA 분자 사이에서 일어날 때 상동적 재조합
⋅재조합 방법은 DNA 분자들 간에 유전물질이 상호 교환되어 재배열되기 때문에 유전적 다양성
이 증가함
⋅재조합 과정은 대개 DNA 복제 또는 복구과정과 밀접히 관련됨
1) 재조합 모델
⋅재조합의 종류 -일반적 재조합(상동성 재조합, homologous genetic recombination)
→ 상동성 서열을 갖는 두 DNA 사이에서 유전물질이 교환되어, 유전적 다양성을 만들어 냄.
두 가닥에 모 두 이상이 생겼을 때, DNA를 복구하는 기작임
-위치특이성 재조합(site-specific recombination)
→ 재조합 효소가 특정 위치를 인식하고 끊어서 새로운 DNA 조각과 이어줌 -유전체 전이과정
(transposition)
→ 자유롭게 이동하는 전이인자(transposon)에 의해 염색체와 염색체 내부 또는 그 사이에
DNA 조각의 이 동과 끼워 넣기를 주도하는 과정
2) DNA의 제한과 변형
⋅대략 4~8개 정도의 특정 염기서열을 가진 DNA 내부를 전달하는 제한효소(restriction
enzyme) 또는 제한적 내부가수분해효소(restriction endonuclease)가 있음
(예) EcoRI 효소 : GATTC와 같이 이중나선의 중앙측을 기준으로 G와 A사이를 절단함
⋅일부 세균들은 이미 제한효소를 갖고 있으며, 이 때 세균은 자신의 DNA 상에 있는 특정 염기
를 메틸화하여 자신의 제한효소 공격으로부터 보호함 → DNA 변형(modification)
⋅제한효소 : 특정 위치의 이중나선 DNA를 절단하는 EcoRI, BamHI 등이 있음
3) 유전공학 Genetic Engineering 또는 재조합 DNA 기술
(1) 제한효소에 의한 특정 DNA 자르기
⋅제한효소들은 특정한 염기 서열을 인식하여 DNA가 끊어지게 함
⋅보통 4~8개까지의 특정 염기서열을 인식하는 제한효소는 DNA를 뭉뚝하게(blunt) 자르거나 한
쪽 가닥이 단 일가닥이 되게(sticky-end) 자름
(2) DNA 연결효소(ligase)
⋅서로 다른 두 DNA 단편을 단단하게 연결시키는 효소
⋅대개 단편으로 끊어진 인산디에스테르 결합을 이어줌
(3) 플라스미드 벡터(plasmid vector)
⋅자연적인 플라스미드를 인공적으로 재조합시켜 만든 플라스미드 백터가 유전공학 기술에서 유
전자를 형질전 환시키는데 쓰임
⋅재조합된 플라스미드 : 스크리닝에 편리한 항생제 내성 유전자가 있고, 그 유전자 내부에 특정
제한효소 인식 DNA 서열에 넣거나, 외부 유전자를 발현시키는 데 좋은 프로모터 DNA를 연결
시키게 함
⋅플라스키더 벡터 DNA는 크기가 비교적 작고, 삽입되는 외부 유전자도 작음
(4) cDNA 합성
⋅인트론 부분이 빠져서 엑손 부분만이 정보를 가진 DNA를 제조하므로 이를 클로닝하여 유용
유전자를 발현 시키는데 좋음
(5) PCR(polymerase chain reaction) 기술
⋅특정 부위의 DNA를 반복적으로 복제시키는 방법으로서 아주 소량의 유전물질을 다량 합성할
수 있음
⋅특정 염기서열의 양쪽 프라이머 사이의 염기서열을 정확히 다량 복제하므로 질병의 진단 등에
유용함
(6) 클로닝(cloning)
⋅플라스미드 벡터 또는 viral 벡터 등에 외부 DNA를 동일한 제한효소로 끊어서 삽입시켜 재조
합 DNA를 만들 고, 숙주세포에 물리적 힘으로 집어넣어 숙주세포를 형질 전환시킴
⋅형질 전환된 세포만 찾아내어 배양시킨 세포(클론)을 얻는 과정
| ※ 클론 : 전체 또는 일부 유전자를 공통으로 보유하고 발현하는 같은 종 또는 개체세포들을 말함 ※ 클로닝 : 클론세포 또는 개체를 얻는 과정을 의미하며, 유전공학 기술에서 외래 유전자를 함유한 클론생 물체를 얻는 과정이 중요함 |
5. 전사과정
⋅전사과정은 DNA의 정보가 RNA 정보로 전환되는 과정으로서, DNA 복제와 유사하게 세 가지
구성요소가 필 요함
-DNA 주형가닥 : RNA 가닥을 만드는데 DNA 가닥이 필요함
-RNA 합성 기질 : ATP, GTP, CTP, UTP가 필요함
-전사기구 : RNA 중합을 촉매하는 단백질로 이루어짐
⋅전사단위(transcription unit)는 RNA로 암호화되는 부분의 DNA 조각으로서 전사에 필요한
염기서열을 말함
⋅프로모터, RNA 암호 부위, 종결인자 등 세가지로 구성됨
-프로모터 : 전사기구가 인식하여 결합하는 DNA 서열, 주형가닥, 전사방향을 정함
-RNA 암호 부위 : RNA 분자가 만들어지는 DNA 상의 염기서열
-종결인자(terminator) : 전사가 끝나는 지역
⋅전사결과, 세포들은 mRNA, tRNA, rRNA 등을 얻음 그 밖에도 조절 또는 촉매 역할을 하는
miRNA 등 작은 크기의 RNA들이 많이 알려져 있음
⋅RNA 합성은 DNA 합성과 달리 프라이머가 필요 없이 리보뉴클레오시드삼인산(NTP)으로부터
합성되기 시작 함
⋅신장 중에는 RNA 분자의 3‘-OH에 NTP가 하나씩 추가됨
⋅두 번째 결합하는 RNA 뉴클레오티드는 바로 앞 뉴클레오티드늬 3‘-OH에 인산디에스테르 결
합으로 연결되 면서 pyrophosphate(PPi)가 떨어져나감
⋅이렇게 합성된 RNA 가닥은 DNA 가닥 중 주형가닥과 상보적이며, 역평행 관계임
1) RNA 중합효소
(1) 세균의 RNA 중합효소(RNA polymerase)
⋅세균은 한 종류의 RNA 중합효소로 만든 RNA를 합성함
⋅RNA 중합효소의 핵심효소는 α-단위체 2개, β-단위체 및 β‘-단위체, ω-단위체로 구성되어
RNA 분자의 신 장을 촉매하고, σ(시그마) 단위체까지 결합된 완전효소는 프로모터를 찾아 결
합시켜 전사를 시작하게 함
(2) 진핵세포의 RNA 중합효소
⋅세 가지 서로 다른 RNA 중합효소들이 서로 다른 종류의 RNA를 합성함
-RNA 중합효소 I (RNA pol I) : pre-rRNA -RNA 중합효소 II (RNA pol II) : mRNA 전구
체, snRNA, miRNA, snoRNA 전사 -RNA 중합효소 III (RNA pol III) : tRNA, 5S rRNA,
miRNA, snRNA 전사
2) 원핵세포의 mRNA 합성과정
(개시, 신장, 종결단계)
(1) 개시단계
⋅RNA합성을 시작하는데 필요한 4가지
① 프로모터 인식
② 전사버블 형성
③ rNTP를 이용하여 첫 번째 뉴클레오티드 결합
④ 프로모터에서 σ-단위체의 벗어남과 전사기구가 프로모터를 떠나는 단계까지를 말함
⋅프로모터는 전사가 시작되는 곳으로서 DNA 두 가닥 중 어느 가닥이 읽혀지고 있으며, 어느
방향으로 중합효소가 움직이는 것인가를 알려줌
⋅세균의 프로모터들은 대부분 RNA 암호화지역 가까이 있으며, 세균들의 여러 유전자에는 공통
적인 염기서열 이 있음
⋅그 위치에 따라 지적되는 공통염기서열은 -10 위치의 5‘-TATAAT-3'으로 표시되며, 이를 보통
TATA 박스 라함
⋅또 다른 위치의 공통염기서열로는 -35위치에 TTGACA 서열이 있음
-RNA 완전중합효소는 NTP의 염기가 시작점 DNA에 있는 상보적 염기와 쌍을 이루게 함
-프로모터에 결합된 채 9~12개 뉴클레오티드 길이의 전사체가 만들어지면 → σ-단위체가 프로
모터 지역에서 떨어져 나감 → 핵심효소만 남아 → 계속 신장단계로 들어감
(2) 신장단계
⋅시그마(σ) 단백질이 완전중합효소에서 떨어져 나오면 → RNA 중합효소는 핵심효소(core)로 바
뀜 → 프로모터 지역을 떠나 아래쪽으로 내려가며 RNA를 신장시킴
⋅핵심효소가 전사를 행하면서 앞쪽 방향으로 내려가면 뒤쪽에서는 일시적으로 주형가닥과 염기
짝을 형성 → 붙어 있던 RNA 전사체는 수소결합이 끊어짐 → 대신 주형 DNA 가닥은 비주형
DNA 가닥과 다시 염기쌍을 이루어 원래 DNA 형태로 돌아감
⋅전사는 중간에 일시중지되었다가 계속 진행하기도 함
(3) 종결단계
⋅종결인자부분까지 모두 전사되면 중합효소는 RNA 합성을 끝내고, 합성된 RNA 전사체가 방출
되며 RNA 중합효소도 DNA 주형에서 떨어져 나옴으로써 전사는 종결됨
6. 유전암호와 단백질 합성
1) 단백질 합성 준비과정 ⋅단백질 합성은 세포질에 있는 리보좀에서 일어남
⋅20개 아미노산은 각각의 아미노아실-tRNA 합성효소에 의해 각 해상 tRNA에 결합됨
⋅이 효소는 tRNA의 안티코돈과 그에 해당하는 아미노산의 R-그룹들을 식별하여 특정 아미노산
을 3‘-OH에 결합시킴
⋅피로인산(PPi)이 가수분해되어 생성되는 에너지는 아미노아실-tRNA가 계속 합성되는 방향으로

| ※ 핵심효소 : 세균의 중합효소는 ααββ'ω 단위체들이 결합되어 RNA 합성의 신장단계를 담당 ※ 완전효소 : 핵심효소에 σ(시그마)-단위체가 결합되면 전사시 프로모터를 인식 |
2) 코돈과 안티코돈
(1) 코돈(codon)
⋅한 아미노산은 실제로 3개의 염기 그룹, 즉 코돈(codon)이라는 3중 암호마다 특정 아미노산으로 읽힘
⋅대부분의 생물체들은 ‘공통 유전암호’를 갖고 있음
⋅코돈의 각 뉴클레오티드는 4개 염기(A, G, C, U) 중 하나를 가질 수 있음 → 64의 코돈 가능
⋅이 중에서 종결 코돈 3개를 제외하면 61개의 전사 코돈이 20가지의 아미노산을 암호화 할 수 있음
(2) 안티코돈(Anti-codon)
⋅tRNA 유전자가 전사 후 몇몇 기본 염기에 구조가 변형된 염기가 나타남
⋅모든 tRNA의 구조는 매우 유사하고, 일부 내부에 상보적으로 수소결합으로 클로버 잎 모양을 함
⋅안티코돈에는 mRNA의 코돈과 상보적으로 결합하는 3개의 염기의 안티코돈이 있음
(3) 와블현상
⋅와블(wobble; 흔들림) 현상 : 코돈의 세 번째 염기와 tRNA 상에 있는 안티코돈 첫 번째 염기
사이의 상호결 합은 약한 상태
⋅그래서 어떤 안티코돈들은 1개 이상의 mRNA 코돈과 짝을 이룸
⋅퇴행성 유전 암호(degenerate code) : 두 개 이상의 코돈이 동일한 한 개의 아미노산을 암호
화할 수 있는 코돈의 성질
① 유전암호에 유연성
② mRNA상의 코돈과 tRNA상의 안티코돈 사이의 결합이 약해서 단백질 합성이 일어나고 있을
때, tRNA를 재빨리 mRNA로부터 분리시켜 단백질 합성 속도를 빨라지게 함
③ 코돈의 세 번째 염기에 해당하는 DNA 상의 염기에 돌연변이가 오더라도 변경된 단백질로 만
들어질 위험을 줄임
(4) 개시코돈과 정지코돈
⋅mRNA 상에서 개시코돈 : AUG(중간에 나온 AUG 코돈은 일반적인 아미노산 메티오닌으로 읽힘)
⋅종결코돈(stop codon) : UAG, UGA, UAA(맞는 아미노아실-tRNA가 없어서 리보좀으로 아미
노산이 들어오지 못하여 종결됨)
⋅개시코돈(AUG) -ORF(Open Reading Frame) -정지코돈의 배열이 이루어짐
⋅ORF : 개시코돈으로부터 종결코돈 바로 앞까지의 중복되지 않는 코돈 서열
3) 단백질 합성
⋅원핵세포의 단백질 합성과 변형에는 100개 이상의 단백질과 여러 종류의 RNA들이 관여함
⋅원핵세포의 단백질 합성과정은 세 단계로 나눌수 있음
⋅진핵세포들의 단백질 합성과정도 원핵세포와 유사함
⋅개시단계 - 신장단계 - 종결과정으로 이루어져 있음
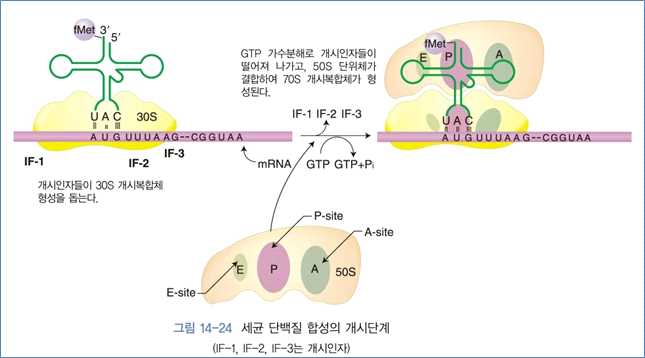
(1) 개시단계
⋅mRNA와 개시인자(IF), 30S 리보좀 단위체들이 GT, N-formylmethionin-tRNAfMet 등과 함
께 30S-개시복합 체를 형성
⋅이 때, mRNA상의 5‘-말단 쪽에 있는 SD(shine-dalgarno) 염기서열과 30S 리보좀의 16S
rRNA 사이에 일시적인 염기쌍 결합으로 복합체가 형성
(2) 신장단계
⋅70S 개시복합체가 형성되고 나서 두 번째 코돈에 맞는 아미노산을 갖고 있는 아미노아실
-tRNA, 신장인자들이 참여함
① EF-Tu가 리보좀의 A-위치에 아미노아실-tRNA를 위치시킴. P자리에는 이미 fMet-tRNA가
있음
② 첫 아미노산인 fMet의 카르복실기(COOH)와 두 번째 아미노산의 아미노기(-NH 2) 사이에 펩
티드 결합 → 2개 아미노산을 가진 디펩티딜-tRNA가 리보좀의 A-위치에 생성
③ 아미노산 fMet을 잃은 P-위치에 남아있던 tRNA는 리보좀에서 떨어져 나가고, 70S 리보좀은
mRNA의 3‘-쪽으로 한 코돈만큼 이동
④ 신장인자 EF-G가 mRNA와 디펩티딜-tRNA를 좌측으로 한 코돈 간격만큼 이동시킴 → 디펩
피딜-tRNA가 P 자리로 이동되고 탈아실화된 첫 번째 tRNA를 밖으로 내보냄
⑤ A 자리는 비어있게 되므로 세 번째 새로운 아미노아실-tRNA가 들어와 펩티드 결합이 반복됨
⋅GTP가 가수분해되어 에너지로 쓰임
⋅즉, 아미노산 하나가 연결될 때마다 GTP 두 개가 하나는 아미노산 운반(아미노아실-tRNA 형
성에 쓰임), 다 른 하나는 A 위치에서 P 위치의 자리 옮김에 쓰임
(3) 종결과정
⋅리보좀 A-위치에 종결코돈인 UAG, UGA, UAA 중 하나가 들어오면, 상보적 안티코돈을 가진
아미노아실 -tRNA가 존재하지 않아서 아미노산을 가진 tRNA가 들어오지 않게 됨
⋅대신 그 자리에 방출인자(RF 1)가 자리하여 펩티드전이효소를 활성화시켜서 tRNA와 단백질
간에 생긴 에스테르 결합이 가수분해되어 서로 떨어지게 함


<확인문제>
문제 1. RNA 전사과정에서 아미노산을 분석 및 해석하기 위해서는 코돈(codon)이 필요하다.
mRNA상에서 복제를 시작하는 단계인 개시코돈은 다음 중 무엇인가? [2016년_9번]
정답 ③ ① UAG ② UGA ③ AUG ④ AGU
문제 2. 유전암호 코돈(codon)에 대한 설명으로 옳지 않은 것은? [2016년_32번]
① 대부분의 생물체들은 공통의 유전암호를 갖고 있다.
② 하나의 아미노산을 지정하는 3개의 염기로 된 서열이다.
③ 한 아미노산이 한 개 이상의 코돈을 가질 수 있다.
정답 ④ ④ 코돈은 3‘ → 5’ 방향으로 읽힌다.
문제 3. 64가지 가능한 코돈 중에서 그에 맞는 아미노아실-tRNA가 없어서 리보좀으로 아미노산이 들어
오지 못하는 코돈을 종결코돈이라고 한다. 다음 중 종결코돈이 아닌 것은? [2017년_26번]
정답 ④ ① UAG ② UGA ③ UAA ④ AUG
문제 4. 64가지 가능한 코돈 중에서 그에 맞는 아미노아실-tRNA가 없어서 리보좀으로 아미노산이 들어
오지 못하는 코돈을 종결코돈 이라고 한다. 다음 중 종결코돈으로만 나열된 것은? [2018년_18번]
① UAA, UGU, UAG
② UAA, UGA, UAG
③ UAA, UGA, UGG
정답 ② ④ UUC, UGA, UAG
문제 5. 유전암호 코돈(codon)에 대한 설명으로 옳은 것은? [2017년_32번]
① 코돈의 뉴클레오티드는 3개 임기를 가지므로 3x3x3=27개의 코돈이 가능하다.
② 하나의 아미노산을 지정하는 3개의 염기로 된 서열이다.
③ 한 개의 코돈은 반드시 한 개의 아미노산만 암호화할 수 있다.
정답 ② ④ 코돈은 3' → 5' 방향으로 읽힌다.
문제 6. 유전암호 코돈(codon)에 대한 설명으로 옳은 것은? [2018년_20번]
① 코돈의 뉴클레오티드는 4개 염기를 가지므로 4 x 4 x 4 = 64개의 코돈이 가능하다.
② 하나의 아미노산을 지정하는 4개의 임기로 된 서열이다.
③ 한 개의 코돈은 반드시 한 개의 아미노산만 암호화할 수 있다.
정답 ① ④ 코돈은 3' → 5' 방향으로 읽힌다.
문제 7. 다음 중 단백질 합성과정에서 tRNA와 결합하는 것으로 볼 수 있는 것은 무엇인가? [2018년_35번]
① AMP ② 펩티드
정답 ③ ③ 아미노산 ④ 안티코돈
문제 8. 다음 중 DNA 복제에 필요한 물질로 옳은 것은?
㉮ RNA 중합효소 ㉯ RNA 프라이머 ㉰ 제한효소 ㉱ DNA 복제효소 I, II, III
정답 ③ ① ㉮㉯㉰㉱ ② ㉮㉰ ③ ㉯㉱ ④ ㉮㉯㉰
문제 9. 다음 중 DNA 전사과정에 필요한 인자로 옳은 것은?
㉮ DNA 주형가닥 ㉯ 전사기구 ㉰ GTP㉱ rRNA
정답 ④ ① ㉮㉯㉰㉱ ② ㉮㉰ ③ ㉯㉱ ④ ㉮㉯㉰
문제10. 다음 중 DNA 복제에 대한 설명 중 옳지 않은 것은?
① 원핵생물 DNA는 양쪽 방향성의 복제를 한다.
② 진핵생물 DNA는 복제개시점이 오로지 하나만 존재한다.
③ 지연가닥은 오카자키조각의 형성으로 완성된다.
정답 ② ④ DNA 복제는 2가닥 모두가 복제시 주형가닥이 된다.
※ 부족하지만 글의 내용이 도움이 조금이라도 되셨다면, 단 1초만 부탁드려도 될까요? 로그인이 필요없는 하트♥(공감) 눌러서 블로그 운영에 힘을 부탁드립니다. 그럼 오늘도 행복한 하루 되십시오^^
'생화학' 카테고리의 다른 글
| 식품, 영양 관련 쉽게 정리한 생화학 핵심 요점 요약 정리 15. 핵산의 구조와 기능 (0) | 2022.05.14 |
|---|---|
| 식품, 영양 관련 쉽게 정리한 생화학 핵심 요점 요약 정리 14. 아미노산 및 질소대사(2) (0) | 2022.05.13 |
| 식품, 영양 관련 쉽게 정리한 생화학 핵심 요점 요약 정리 13. 아미노산 및 질소대사(1) (0) | 2022.05.12 |
| 식품, 영양 관련 쉽게 정리한 생화학 핵심 요점 요약 정리 12. 지질 대사(2) & 아미노산 및 질소대사(1) (0) | 2022.05.11 |
| 식품, 영양 관련 쉽게 정리한 생화학 핵심 요점 요약 정리 11. 지질대사 (0) | 2022.05.10 |